

1. Introduction
2. Quercetin as a Biological Regulator in HCC Models
3. Model-Dependent Antiproliferative Effects
4. Redox Regulation and p53-Related Apoptosis
5. Survival Signaling and Cell-Cycle Control
6. Metabolic Vulnerability Through FASN and Lipid Biosynthesis
7. Experimental Chemosensitization and Drug-Combination Evidence
8. In Vivo Evidence and Barriers to Translation
9. Future Perspective
10. Conclusion
1. Introduction
간세포암(hepatocellular carcinoma, HCC)은 원발성 간암의 대표적 형태이며, 전 세계 암 사망 부담에서 중요한 위치를 차지한다. 2022년 GLOBOCAN 자료는 간암이 여전히 높은 사망 부담을 보이는 암종임을 제시하였고[1], HCC는 원발성 간암의 대부분을 차지하는 질환으로 만성 간염, 간경변, 대사성 간질환 및 간 내 염증 환경과 밀접하게 연결된다[2]. 최근 HCC 치료는 감시, 절제, 간이식, 국소소작술, 경동맥 치료, 표적치료 및 면역치료를 포함하는 다층적 전략으로 발전하였다[3,4]. 그러나 HCC는 종양 자체의 분자적 이질성뿐 아니라 환자의 기저 간기능, 간경변 동반 여부, 면역미세환경, 재발 위험이 함께 치료 반응을 결정한다는 점에서 단일 항암전략으로 설명하기 어렵다[2,3]. 2025년 AASLD critical update가 제시하듯이 HCC 치료 권고는 빠르게 갱신되고 있으며, 이는 현재 치료법의 발전과 동시에 남아 있는 임상적 불확실성을 반영한다[5].
이러한 배경에서 식물 유래 생리활성물질(phytochemical)은 HCC의 표준치료를 대체하는 물질이 아니라, 암세포의 취약성을 이해하고 보조적 치료전략을 탐색하기 위한 전임상 연구 소재로 다루어져 왔다. 플라보노이드 계열 물질은 다양한 식품에 존재하며 산화스트레스, 염증반응, 세포 성장 신호 및 세포사멸 관련 경로에 영향을 줄 수 있다[6,7]. 그러나 phytochemical 연구에서 흔히 발생하는 문제는 세포주 수준의 긍정적 결과가 곧바로 임상적 치료효과처럼 과장되는 것이다. 따라서 quercetin의 HCC 관련 연구도 “항암효과가 있다”는 단순 결론보다, 어떤 모델에서 어떤 조건으로 어떤 생물학적 축이 변화하였는지를 구분하여 해석해야 한다.
Quercetin은 대표적인 플라보놀 계열 화합물로, HCC 관련 전임상 모델에서 세포 생존율 감소, apoptosis 유도, 세포주기 변화, 생존신호 억제 및 지방산 합성 저해와 연결되어 보고되었다. 본 논문에서 제시하는 중심 논제는 quercetin이 HCC 치료제로 확립되었다는 것이 아니라, HCC 관련 전임상 모델에서 redox-p53/apoptosis-survival signaling-lipid metabolism 축을 동시에 흔들 수 있는 다중표적 후보물질이라는 점이다. 동시에 이 효과는 HepG2 중심 모델 의존성이 크고, 세포주 이질성, 생체이용률, 약동학, 정상 간세포 선택성, 면역미세환경 반영 부족이라는 한계 때문에 임상적 의미로 바로 확장할 수 없다. 따라서 본 리뷰는 quercetin의 가능성과 한계를 같은 비중으로 해석하여, 향후 검증되어야 할 연구 방향을 제시하고자 한다.
2. Quercetin as a Biological Regulator in HCC Models
Quercetin을 HCC 연구에서 의미 있게 다루려면 먼저 이 물질을 “단일 표적 항암제”가 아니라 “세포 상태에 따라 여러 신호망에 동시에 관여할 수 있는 생리활성 조절자”로 보는 것이 적절하다. Quercetin은 항산화 물질로 소개되는 경우가 많지만, 암세포에서 산화환원 균형은 단순한 보호 또는 손상 반응으로 나뉘지 않는다. 암세포는 정상세포와 달리 높은 대사활성, 미토콘드리아 기능 변화, 성장신호 활성화로 인해 ROS 의존적 생존과 ROS 유발 손상 사이의 좁은 균형을 유지한다[8,9]. 따라서 quercetin이 ROS를 낮추거나 특정 조건에서 산화환원 스트레스를 조절한다는 결과는 단순히 “항산화 작용”으로 끝나는 것이 아니라, 암세포가 유지하던 redox-dependent survival window를 변화시키는 현상으로 해석해야 한다.
HCC에서는 p53 경로의 상태도 quercetin 반응을 해석하는 데 중요하다. p53 신호는 DNA 손상 반응, 세포주기 정지, apoptosis, 대사 조절에 관여하며 HCC 발생 및 진행에서 핵심적인 종양억제축으로 다루어진다[10,11]. Quercetin 관련 HCC 연구에서 HepG2가 반복적으로 사용된 이유도 이 세포주가 p53 관련 반응을 관찰하기 쉬운 모델이기 때문이다. 그러나 이 장점은 동시에 한계가 된다. HepG2는 성인 HCC에서 유래한 전형적 간세포암 세포주라기보다 hepatoblastoma-derived cell line으로 재해석되어야 한다는 지적이 있으며[12], HepG2를 일반 HCC의 대표 모델처럼 사용하는 관행 자체도 비판적으로 검토되어 왔다[13]. 따라서 quercetin의 HepG2 효과는 HCC 전반의 효과라기보다 특정 간암 관련 세포 모델에서 관찰된 redox 및 p53/apoptosis 의존성 반응으로 제한하여 해석해야 한다.
이와 같은 관점은 이미 보고된 quercetin-HCC 종합문헌과도 연결된다. 기존 리뷰들은 quercetin이 hepatocarcinoma 모델에서 세포사멸, 세포주기 정지, 산화스트레스 조절 및 대사 조절에 관여할 수 있음을 정리하였다[14,15]. 그러나 본 논문은 기존 결과를 넓게 나열하기보다, HCC 전임상 모델에서 quercetin 반응이 redox 조절, p53/BAX-caspase 축, PI3K/Akt/ERK 생존신호, 세포주기, FASN 기반 지질대사 취약성이라는 연결망으로 해석될 수 있는지에 초점을 둔다.
3. Model-Dependent Antiproliferative Effects
Quercetin의 HCC 관련 항증식 효과는 HepG2 세포에서 비교적 일관되게 관찰되었다(Table 1). Granado-Serrano 등은 HepG2 세포에 quercetin을 처리했을 때 노출 시간이 충분할 경우 세포 생존율이 감소하고 apoptosis가 증가하며, caspase-3 및 caspase-9 활성화가 동반된다고 보고하였다[16]. 이 연구에서 짧은 노출 시간은 충분한 세포사멸 반응을 유도하지 못했으므로, quercetin 반응은 농도뿐 아니라 처리 시간에 강하게 의존한다. 이 지점은 매우 중요하다. 세포주 실험에서 특정 농도의 효과를 제시하더라도, 노출 시간, 세포 밀도, 배지 조건, 혈청 존재 여부, quercetin 용해 방식이 달라지면 결과가 달라질 수 있다. 따라서 IC50 또는 특정 농도 반응을 절대값처럼 해석하는 것은 부정확하다.
Table 1.
Experimental evidence and interpretation of quercetin activity in HCC-related models
| Model | Main observation | Logical interpretation | Major limitation |
| HepG2 | Reduced viability and increased apoptosis after sufficient exposure, with caspase-3/9 activation [16]. | HepG2 provides strong mechanistic evidence for apoptosis-associated growth suppression. | HepG2 does not represent the full molecular diversity of adult HCC. |
|
HepG2 and Huh7 comparison | HepG2 responded to quercetin, whereas Huh7 showed limited response under comparable conditions [17]. | Cell-line background and redox response are central to interpretation. | The difference should not be reduced only to p53 status without additional validation. |
|
Multiple liver cancer cell lines | Dose- and time-dependent suppression was observed across several lines, but sensitivity differed [18]. | Quercetin may affect multiple models, but response heterogeneity is substantial. | Predictive biomarkers remain unclear. |
|
HepG2 xenograft | Delayed tumor growth, increased necrosis, improved survival, and reduced cyclin D1 staining were reported [19]. | In vivo tumor-growth delay supports preclinical potential. | Subcutaneous nude mouse models do not reproduce immune and cirrhotic HCC environments. |
Jeon et al.(2019)은 HepG2와 Huh7을 비교하여 quercetin 반응의 세포주 의존성을 더 명확하게 제시하였다[17]. 이 연구에서 quercetin은 HepG2 세포의 증식을 억제하고 intracellular ROS 수준을 낮추었지만, Huh7 세포에서는 동일한 수준의 항증식 반응이 제한적이었다[17]. 이는 quercetin의 효과가 HCC라는 병명 자체보다 세포주의 분자적 배경에 의존할 가능성을 보여준다. Huh7은 mutant p53 배경을 가진 세포주로 자주 다루어지므로, HepG2 중심의 p53 관련 반응을 Huh7 또는 다른 HCC 모델로 그대로 확장하는 데에는 주의가 필요하다. 다만 Jeon 등의 결과는 quercetin의 항증식 효과가 p53 변화만으로 설명되지 않고 ROS 조절과 밀접하게 연결될 수 있음을 함께 보여준다[17].
다중 세포주 자료도 같은 결론을 지지한다. Hisaka 등은 13종의 liver cancer cell line에서 quercetin의 농도 및 시간 의존적 증식 억제를 분석하고, 일부 세포주에서 apoptosis와 cell-cycle arrest가 동반된다고 보고하였다[18]. 다중 세포주 자료는 quercetin의 효과가 HepG2에만 국한되지 않을 수 있음을 보여주는 장점이 있지만, 동시에 세포주별 감수성 차이가 크다는 사실도 드러낸다. 그러므로 quercetin을 “HCC 세포에 보편적으로 작용하는 물질”로 서술하기보다 “특정 분자적 조건을 가진 HCC 관련 모델에서 증식 억제 신호를 유도할 수 있는 물질”로 서술하는 것이 더 정확하다.
4. Redox Regulation and p53-Related Apoptosis
Quercetin의 HCC 전임상 효과를 해석할 때 가장 먼저 정리해야 할 축은 ROS 조절이다. 암세포에서 ROS는 DNA 손상과 세포사멸을 유도할 수 있지만, 동시에 증식, 침윤, 생존신호, 대사 재편성을 돕는 신호분자로도 작용한다[8,9]. 따라서 quercetin이 ROS를 변화시킨다는 결과는 “ROS가 증가했다” 또는 “ROS가 감소했다”는 단편적 설명으로 끝나서는 안 된다. 핵심은 quercetin이 해당 세포가 의존하던 산화환원 상태를 변동시켜 생존신호와 세포사멸 신호의 균형을 바꾸는가이다.
Jeon et al.(2019)은 HepG2에서 quercetin의 항증식 효과가 intracellular ROS 감소와 관련되며, H2O2 보충을 통해 quercetin 효과가 약화되는 현상을 제시하였다[17]. 이 결과는 quercetin의 항증식 반응에서 ROS 감소가 단순한 동반 현상이 아니라 기능적으로 연결된 과정일 가능성을 뒷받침한다. 다만 이 연구에서도 모든 HCC 세포가 동일하게 반응하지 않았으므로, ROS 조절은 quercetin의 보편적 항암기전이라기보다 특정 세포 상태에서 작동하는 upstream interpretive axis로 보는 것이 타당하다.
ROS 조절 다음에는 p53/BAX-caspase 축이 연결된다. Granado-Serrano 등은 HepG2에서 quercetin이 caspase-3 및 caspase-9 활성화, Bcl-2 family 조절, Akt 및 ERK 생존신호 억제와 함께 apoptosis를 유도한다고 보고하였다[16]. Maurya와 Vinayak은 quercetin이 PI3K, PKC, COX-2를 낮추고 p53 및 BAX 발현을 증가시킨다고 제시하였다[20]. 최근 Guan 등도 HepG2 세포에서 quercetin이 YY1 억제를 통해 p53 경로와 연결될 수 있음을 보고하였다[21]. 이러한 자료를 종합하면, quercetin의 apoptosis 유도는 단일 경로가 아니라 redox 변화, p53/BAX 증가, caspase-3/9 활성화, 생존신호 약화가 함께 나타나는 복합 반응이다.
그러나 p53 축을 지나치게 강조하는 것도 위험하다. Jeon 등의 비교 연구에서 quercetin 반응은 p53 상태만으로 설명되지 않았고, ROS 조절이 더 직접적인 해석축으로 제시되었다[17]. 따라서 quercetin의 HCC 관련 기전은 “p53 유도에 의한 apoptosis”라는 단순 문장보다 “redox 변화가 생존신호와 p53/BAX-caspase 반응을 함께 재배열하는 모델 의존적 apoptosis network”로 해석하는 것이 더 정확하다.
5. Survival Signaling and Cell-Cycle Control
Quercetin은 apoptosis 신호만이 아니라 생존신호 억제와 세포주기 조절에도 관여하는 것으로 보고되었다(Table 2). HepG2 연구에서 PI3K/Akt 및 ERK 경로는 quercetin 처리 후 억제되는 경향을 보였으며[16,20], 이는 apoptosis 감수성을 증가시키는 배경으로 해석될 수 있다. PI3K/Akt와 ERK는 HCC를 포함한 여러 암종에서 세포 생존, 성장, 대사, 약제 저항성과 연결되는 주요 신호축이다. Quercetin이 이 경로를 낮춘다는 관찰은 quercetin이 단순 세포독성 물질이 아니라 survival signaling pressure를 낮추는 조절자로 작용할 수 있음을 시사한다.
Table 2.
Mechanistic axes connecting quercetin exposure to antiproliferative responses
| Mechanistic axis | Representative evidence | Interpretive value | Caution |
| Redox regulation | Intracellular ROS reduction and H2O2 rescue in HepG2 [17]. | Redox state may function as an upstream condition for quercetin sensitivity. | ROS response is context-dependent and cannot be generalized to all HCC models. |
|
p53/BAX-caspase apoptosis | p53/BAX increase, caspase-3/9 activation, and mitochondrial apoptosis-related changes [16,20,21]. | Supports apoptosis as a central downstream outcome in responsive models. | p53 status alone does not fully predict response. |
|
Survival signaling suppression | PI3K/PKC/COX-2 decrease and Akt/ERK inhibition [16,20]. | May lower the survival threshold and increase apoptosis sensitivity. | Direct molecular binding targets remain incompletely defined. |
| Cell-cycle control | Cyclin A, CHK1 and cyclin D1 regulation [17,19,22]. | Suggests growth suppression through both cell death and cell-cycle slowing. | Cell-cycle phase effects vary by model and assay design. |
|
Cell structure and invasion-related proteins | Sp1-associated migration/invasion effects and proteomic changes involving IQGAP1 or beta-tubulin [23,24]. | Extends interpretation beyond viability assays. | Evidence is supportive but not sufficient for clinical invasion claims. |
세포주기 조절도 quercetin 반응의 중요한 하위축이다. Jeon 등은 quercetin 처리 후 cyclin A와 CHK1 변화가 세포주기 조절과 관련될 수 있음을 제시하였다[17]. Zhou 등은 HepG2 세포에서 quercetin이 cyclin D1 발현을 감소시키고 G1 cell-cycle arrest와 연결될 수 있음을 보고하였다[22]. Zhou 등의 in vivo 연구에서도 xenograft 조직에서 cyclin D1 staining 감소가 관찰되었다[19]. 이 결과들은 quercetin이 apoptosis를 유도하는 동시에 세포주기의 진행 속도를 늦추어 증식 억제 효과를 강화할 가능성을 보여준다(Fig. 1).
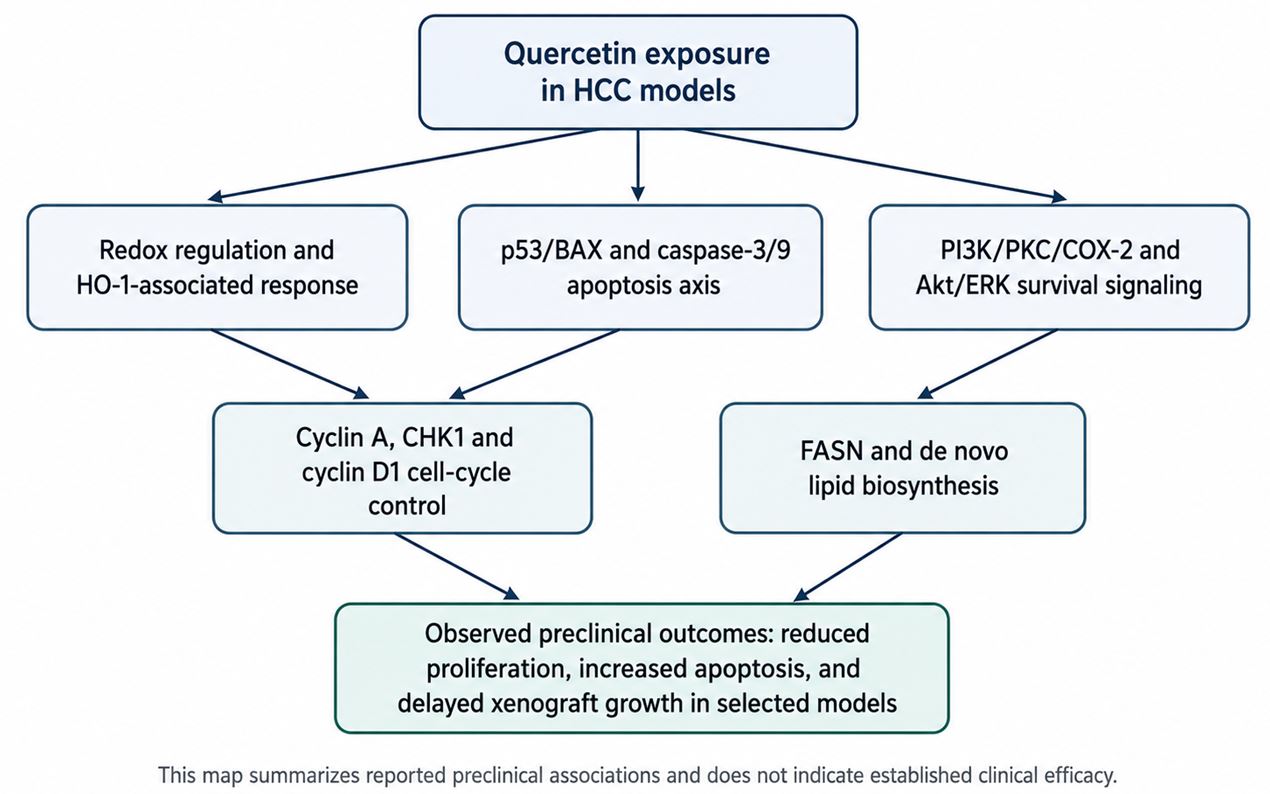
Fig. 1
Mechanistic interpretation of quercetin-associated responses in HCC experimental models. The figure summarizes reported preclinical associations among redox regulation, p53/BAX-caspase signaling, survival signaling, cell-cycle control, lipid metabolism, and observed growth suppression. This schematic does not indicate established clinical efficacy.
또 다른 보조기전으로는 세포골격 및 이동성 관련 단백질 변화가 있다. Zhou 등은 HepG2 proteomic analysis에서 quercetin에 의해 IQGAP1, beta-tubulin과 같은 세포구조 및 이동성 관련 단백질 변화가 나타날 수 있음을 제시하였다[24]. Lee 등은 quercetin이 Sp1 조절을 통해 HepG2 세포의 migration과 invasion 관련 반응을 억제할 수 있다고 보고하였다[23]. 이 자료들은 quercetin 연구를 단순한 세포 생존율 감소에서 세포주기, 세포구조, 침윤성 조절로 확장할 수 있음을 보여주지만, 관련 결과는 아직 HCC 임상 침윤성을 설명할 만큼 충분히 검증된 것은 아니다.
6. Metabolic Vulnerability Through FASN and Lipid Biosynthesis
HCC는 단순히 증식 신호가 증가한 암이 아니라, 간세포의 대사적 특성과 암세포의 생합성 요구가 복합적으로 얽힌 질환이다. 이 점에서 FASN(fatty acid synthase)은 quercetin 연구에서 중요한 해석축이 된다. FASN은 de novo fatty acid synthesis의 핵심 효소로, 암세포의 막 생합성, 에너지 저장, 신호지질 생산, 성장 유지와 연결된다. HCC에서도 FASN 및 관련 지방산 생합성 경로는 발암, 진행, 예후 및 치료표적 가능성과 관련된 축으로 다루어진다[25,26].
Zhao et al.(2014)은 HepG2 세포에서 quercetin이 FASN 활성을 낮추고, 지방산 생합성 억제를 통해 apoptosis를 유도할 수 있음을 보고하였다[27]. 특히 palmitic acid rescue 실험은 FASN 억제가 단순 동반현상이 아니라 quercetin 반응과 기능적으로 연결될 수 있음을 시사한다[27]. 이 결과는 quercetin의 항증식 효과를 redox 및 apoptosis 신호만이 아니라 암세포의 지질대사 취약성으로 확장하여 해석할 수 있게 한다.
다만 FASN 축도 과장해서는 안 된다. Zhao et al.(2014)의 결과는 주로 HepG2 기반이며, 다른 HCC 세포주, 정상 간세포, 3D 배양, organoid, orthotopic model에서 같은 수준으로 검증되었다고 말할 수 없다. 따라서 FASN 억제는 quercetin의 확정적 항암기전이 아니라, HCC의 대사 취약성과 연결될 수 있는 유력한 후보축이다. 향후 연구에서는 quercetin 반응성이 높은 세포주와 낮은 세포주 사이에서 FASN 발현, lipogenic flux, palmitate rescue, SREBP1/ACC/AMPK 관련 변화가 함께 검증되어야 한다(Fig. 1).
7. Experimental Chemosensitization and Drug-Combination Evidence
기존 연구에서 cisplatin과 5-fluorouracil(5-FU)을 HCC의 “1차 화학요법제”처럼 표현한 부분은 적절하지 않다. 현재 HCC 전신치료 환경은 면역치료 및 표적치료 기반으로 빠르게 변화하고 있으며[3,4,5], cisplatin 또는 5-FU 병용 결과를 곧바로 현재 HCC 임상 병용요법으로 해석해서는 안 된다. 이들 연구는 임상 권고가 아니라, quercetin이 세포독성 항암제 반응성을 높일 수 있는지 평가한 전임상 chemosensitization 자료로 제한하는 것이 타당하다(Table 3).
Table 3.
Combination evidence of quercetin in HCC-related models
| Combination | Reported finding | Possible interpretation | Required validation |
| Quercetin plus cisplatin | Stronger growth suppression and apoptosis than either agent alone in HepG2 [28]. | Quercetin may enhance apoptosis and G1 arrest under selected conditions. | PK/PD relevance, hepatocyte toxicity, and clinically relevant dosing. |
| Quercetin plus 5-FU | Enhanced apoptosis or growth suppression in HCC-related models [18,29]. | Supports experimental chemosensitization. | Cell-line-specific response and combination scheduling. |
|
Quercetin and multidrug resistance | FZD7/beta-catenin-associated reversal of multidrug resistance was reported [30]. | Quercetin may interfere with resistance-associated signaling. | Validation in resistant patient-derived models. |
Zhao et al.(2014)은 HepG2 세포에서 quercetin과 cisplatin 병용이 단독 처리보다 성장 억제와 apoptosis를 더 강하게 유도하며, p16 관련 G1 cell-cycle arrest가 병용 반응에 관여할 수 있다고 보고하였다[28]. Dai et al.(2016)은 quercetin이 HCC 모델에서 apoptosis를 유도하고 5-FU의 치료효과를 높일 수 있음을 제시하였다[29]. Hisaka 등도 여러 liver cancer cell line에서 quercetin과 5-FU 병용 효과를 분석하였고, 일부 세포주에서 추가적 또는 상승적 억제가 관찰되었다[18]. 또한 Chen et al. (2018)은 quercetin이 FZD7/beta-catenin 경로를 통해 HCC 세포의 multidrug resistance를 일부 역전시킬 수 있다고 보고하였다[30].
이 자료들은 quercetin이 기존 항암제와 병용될 때 세포사멸 민감도를 높이거나 약제저항성 경로를 약화시킬 가능성을 보여준다. 그러나 임상적으로 의미 있는 병용전략이 되려면 최소 네 가지 조건이 필요하다. 첫째, 병용 농도가 생체 내 도달 가능한 농도인지 확인해야 한다. 둘째, 정상 간세포 또는 간기능 저하 환경에서 독성 선택성이 검증되어야 한다. 셋째, 병용 순서와 노출 시간이 재현성 있게 정리되어야 한다. 넷째, 반응성 예측 바이오마커가 필요하다. 그러므로 현재 단계에서 quercetin 병용효과는 임상 병용요법이 아니라 실험적 sensitizer 가능성으로 제한하여 서술해야 한다.
8. In Vivo Evidence and Barriers to Translation
In vivo 자료는 quercetin의 전임상 가능성을 보완하지만, 현재 수준으로 임상 효능을 주장하기에는 부족하다. Zhou et al.(2014)은 BALB/c nude mouse에 HepG2 세포를 이식한 후 quercetin을 복강투여하였고, 대조군보다 종양 성장 지연, 생존율 증가, 종양 괴사 증가 및 cyclin D1 염색 감소를 보고하였다[19]. 이 결과는 quercetin이 생체 내에서도 종양 성장 관련 지표를 변화시킬 수 있음을 시사한다. Ji et al.(2019)은 quercetin이 HCC 모델에서 autophagy stimulation과 apoptosis induction을 연결할 가능성을 제시하였다[31]. 이러한 연구들은 quercetin이 세포주 실험에만 머무르지 않고 생체 내 종양 환경에서도 일부 반응을 보일 수 있음을 뒷받침한다(Table 4).
Table 4.
Translational barriers and future validation requirements
그러나 현재 in vivo 근거는 사람 HCC의 복잡성을 충분히 반영하지 못한다. Nude mouse는 T cell 결핍 특성이 있어 면역미세환경을 평가하기 어렵고, subcutaneous xenograft는 간경변, 만성염증, portal circulation, 간 내 대사환경, 면역치료 반응성을 재현하지 못한다. HCC는 간조직 자체의 염증, 섬유화, 재생, 혈관화, 면역세포 구성과 강하게 연결되므로[2,3], 피하 HepG2 xenograft에서 종양 성장이 늦어졌다는 결과를 사람 HCC 치료효과로 해석할 수 없다.
가장 큰 translational barrier는 생체이용률이다. Quercetin은 낮은 수용성, 광범위한 대사, 흡수율 차이, 식이 matrix 및 개인차에 의해 혈중 농도와 조직 노출이 크게 달라진다[32,33]. 최근 인체 중재 연구를 종합한 systematic review도 quercetin bioavailability를 높이기 위한 제형, matrix, co-administration 전략이 계속 연구되고 있음을 보여준다[34]. 따라서 세포주에서 사용한 µM 수준의 quercetin 농도를 사람 혈중 또는 간조직 내 유효농도와 직접 연결하는 것은 근거가 부족하다. 이 문제를 해결하기 위해 nanoformulation, encapsulation, targeted delivery와 같은 전략이 제안되었지만[35], 이러한 전달기술 역시 HCC 치료효과를 입증한 것이 아니라 생체이용률 한계를 극복하기 위한 연구 방향으로 이해해야 한다.
9. Future Perspective
향후 quercetin-HCC 연구는 “quercetin이 HCC에 좋다”는 일반론보다 “어떤 HCC 모델이 quercetin에 반응하는가”라는 질문으로 이동해야 한다. 이를 위해서는 HepG2, Huh7, Hep3B, PLC/PRF/5 등 다양한 세포주에서 p53 status, basal ROS level, FASN expression, PI3K/Akt activity, Wnt/beta-catenin status, drug-resistance phenotype을 함께 비교하는 연구가 필요하다. 특히 quercetin 반응성을 설명할 수 있는 biomarker가 없으면, quercetin은 다중표적 물질이라는 장점에도 불구하고 재현성 낮은 phytochemical 연구로 남을 위험이 있다.
두 번째 방향은 model sophistication이다. 2D 세포주 실험은 기전 탐색에는 유용하지만, HCC의 실제 병태를 설명하기에는 부족하다. 3D spheroid, patient-derived organoid, co-culture system, stellate cell 또는 immune cell 포함 모델, orthotopic HCC model을 이용하면 quercetin이 단순히 세포 증식을 억제하는지, 아니면 간암 미세환경에서 의미 있는 생물학적 변화를 유도하는지 더 엄밀하게 판단할 수 있다. 또한 quercetin 자체와 quercetin metabolite의 효과를 구분해야 한다. 실제 생체 내에서는 parent compound보다 glucuronide, sulfate, methylated metabolite가 중요할 수 있으므로[32,33], 세포주 실험도 대사체 노출 조건을 반영하는 방향으로 확장되어야 한다.
세 번째 방향은 delivery and safety이다. Quercetin의 낮은 생체이용률은 단순한 기술적 문제가 아니라, 세포주 결과를 임상으로 옮길 때 가장 큰 해석 장벽이다[32,33,34,35]. 따라서 단순 quercetin aglycone 처리 실험뿐 아니라, 생체이용률 개선 제형, 간 표적 전달체, 독성 선택성, 정상 간세포 보호 또는 손상 가능성, 기존 HCC 치료제와의 상호작용을 함께 평가해야 한다. 이때 목표는 quercetin을 독립 치료제로 주장하는 것이 아니라, HCC의 redox-metabolic vulnerability를 겨냥하는 보조적 또는 탐색적 전략으로서 타당성을 검증하는 것이다.
10. Conclusion
Quercetin은 HCC 관련 전임상 모델에서 세포 생존율 감소, apoptosis 유도, ROS 조절, PI3K/PKC/COX-2 및 Akt/ERK 생존신호 억제, 세포주기 조절, FASN 기반 지방산 생합성 억제와 연결된 항증식 효과를 보이는 것으로 정리된다. 그러나 이 효과는 “HCC 치료효과”라는 단일 결론으로 환원될 수 없다. 더 타당한 해석은 quercetin이 특정 HCC 관련 모델에서 redox-p53/apoptosis-survival signaling-lipid metabolism 축을 동시에 조절하며, 그 결과 증식 억제와 세포사멸 증가가 나타날 수 있다는 것이다.
현재 근거의 중심은 HepG2 기반 in vitro 연구와 제한적인 xenograft 연구에 놓여 있다. Huh7 및 다중 liver cancer cell line 자료는 세포주별 반응 차이를 보여주며, 이는 quercetin의 효과가 HCC 전체에 보편적으로 적용될 수 없음을 의미한다. 낮은 생체이용률, 약동학적 불확실성, 정상 간세포 선택성 부족, 면역미세환경 반영 부족, 임상시험 부재 역시 핵심 한계이다. 따라서 quercetin은 HCC 치료제로 확립된 물질이 아니라, 반응성 바이오마커, 약물전달, 독성 선택성, 환자유래 모델 및 생체 내 재현성 검증이 필요한 다중표적 전임상 후보물질로 평가하는 것이 가장 타당하다.
