

1. Introduction
2. Materials and Methods
2.1. 시약
2.2. 유산균의 분리
2.3. 유산균의 배양
2.4. 유산균 유래 배양 상징액의 제조
2.5. 항균 활성 측정
2.6. SDS-PAGE와 Silver Staining
3. Results and Discussion
3.1. 유산균의 분리 및 동정
3.2. 동치미 유래 유산균의 항균 활성
3.3. 분자량별 상징액의 항균 활성 분석
3.4. 전기영동을 통한 항균활성 분자량 확인
4. Conclusions
1. Introduction
유산균은 발효 과정에서 당을 분해하면서 유산을 비롯해 과산화수소, 디아세틸, 단쇄지방산, 박테리오신 등 다양한 항균 물질을 생성하며, 이를 통해 유해 미생물의 성장을 억제하고 식품의 향미 및 품질을 개선하는 데 기여한다. 이러한 유산균의 대사산물은 인체에 무해하다는 점에서 미국 식품의약국(FDA)으로부터 GRAS (Generally Recognized As Safe)로 인정받았고, 유럽연합의 QPS (Qualified Presumption of Safety) 기준을 충족하여 식품 내 천연 보존제로 활용되고 있다. 유산균은 장 건강 증진을 비롯한 다양한 생리활성이 보고되면서 probiotics로서 그 중요성이 부각되어 왔다. 특히 부패균 및 식중독 유발균을 억제하는 효과로 인해, 유산균은 미생물 오염이 심하고 신선도 유지가 어려운 식품의 저장 기간을 연장하는 생물학적 보존제로서 높은 잠재력을 가진다[1]. 이에 따라 전통 발효식품으로부터 새로운 유산균을 분리하고 이들의 기능성을 규명하는 연구는 학문적으로 큰 의미를 가진다. 최근 식품은 단순한 영양공급을 넘어서 건강을 증진하는 역할까지 기대되고 있으며, 균형 잡힌 식단은 각종 퇴행성 질환의 발생을 지연시키고 증상을 완화하는 데 도움을 줄 수 있다. probiotics 제품은 다양한 질환의 예방 및 치료 목적으로 활용되고 있으며, 이는 Metchnikoff 박사가 제안한 “유익한 미생물의 섭취가 유해균의 증식을 억제한다”는 이론에 뿌리를 두고 있다[2]. 국제식량농업기구(FAO)와 세계보건기구(WHO)는 probiotics를 “적절한 양으로 섭취할 경우 숙주에게 건강상의 이점을 제공할 수 있는 살아있는 미생물”로 정의하였으며[3], 이들의 기능성은 체외 세포배양 모델에서의 평가를 시작으로, 동물실험과 인체 임상시험을 통해 과학적으로 입증되어야 한다[4]. Probiotics는 항생제 연관 설사 완화, 유당불내증 조절, 면역 기능 향상, 알레르기 예방 등 다양한 건강 효과가 보고되고 있으나, 고혈압 조절, 칸디다증 치료 등 일부 특정 질환에 대해서는 아직 근거가 부족하다는 지적도 있다[5, 6, 7]. 기존에는 probiotics가 주로 유제품을 기반으로 개발되어 왔으나, 최근에는 발효 채소류와 같은 식물성 기원을 활용한 연구도 활발히 진행되고 있다[8]. 채소류는 원래 항산화 물질, 식이섬유, 비타민, 무기질 등이 풍부하며, 여기에 probiotics 기능이 결합될 경우 건강 증진 효과를 더욱 극대화할 수 있다[9]. 자연 상태의 대부분의 식물 표면에는 발효가 진행됨에 따라 유산 생성 미생물이 우점하게 되며, 이 중 Lactobacillus 속이 주요한 비율을 차지하는 것으로 보고되고 있다[10]. 하지만 유산균의 생장과 활성을 결정하는 요인은 산도, 염도, 식물의 종류 등 다양한 환경적 요인이 복합적으로 작용한다[11]. 무를 주원료로 하는 동치미는 한국전통 발효식품으로 청량감과 신선미가 매우 우수하다. 이에 본 연구에서는 동치미로부터 유산균을 분리하고, 분리된 균주의 특성을 평가함으로써 향후 probiotics 및 천연 항균 소재로서의 활용 가능성을 탐색하고자 하였다.
2. Materials and Methods
2.1. 시약
전기영동을 위한 시약으로 acrylamide(30%), Coomassie Brilliant Blue (CBB) R-250, Urea는 TNT Research(전주, 한국)에서 구입하였다. Tris, sodium dodecyl sulfate (SDS), ammonium persulfate (APS), tetramethylethylenediamine (TEMED), glycine 및 sodium chloride는 Amresco (Ohio, USA)의 제품을 사용하였다. 2-mercaptoethanol과 bromophenol blue (BPB)는 Sigma (MO, USA) 제품을, glycerol, acetic acid, methanol은 Daejung Chemicals & metals (Siheung, Korea)에서 구매하여 사용하였다.
2.2. 유산균의 분리
동치미로부터 유산균을 분리하기 위하여 경기도 용인시에 소재한 가정에서 제조된 것을 시료로 이용하였다. 동치미시료에 멸균 증류수를 1:10 비율로 첨가한 후, 약 5분간 혼합하여 충분히 미생물이 용출되도록 혼합하였다. 용출된 시료는 즉시 멸균된 peptone water를 이용하여 10배 계열 희석하였으며, 희석액 100 µL을 0.01% Sodium azide(Sigma, St. Louis, MO, USA)가 포함된 De Man Rogosa-Sharpe(Difco, USA) agar에 도말하였다. 도말된 배지는 37°C에서 호기적 및 혐기적 조건 모두에서 배양하여 유산균 콜로니의 생장을 유도하였다. 콜로니 형성 후, 각각의 배양된 MRS agar에서 모양, 크기 및 색이 다른 콜로니를 선택하였다. 본 실험에서는 37°C에서 혐기 배양된 콜로니를 선택하였고, 이 균은 호기상태에서도 생육이 가능하였다. 선택된 유산균은 API 50 CHL kit (BioMerieux Co., Marcy-I’Etoile, France)을 이용하여 당 이용성을 조사하였다. 즉 단일 콜로니를 접종한 CHL 배지를 각 capsules에 100 µL씩 분주 후 37°C에서 48시간 배양하여 색의 변화를 관찰하였다. 당 발효패턴을 API web software에 입력하여 동정 결과를 확인하였다.
2.3. 유산균의 배양
동치미로부터 분리한 유산균의 배양 상징액에 대한 항균활성은 한국미생물보존센터(KCTC)에서 분양 받은 Escherichia coli 균주(11569, 11587, 11591, 11596, 11600)를 대상으로 평가하였다. 시험에 이용된 E. coli 균주는 Luria-Bertani broth(LB)배지(Difco, USA)에서 호기 또는 혐기 조건하에 배양하였다.
2.4. 유산균 유래 배양 상징액의 제조
동치미로부터 분리한 유산균은 MRS broth에 접종한 후, 37°C 혐기 조건에서 24시간 배양하였다. 배양이 완료된 후, 유산균의 활성을 억제하기 위하여 배양액을 얼음 위에 약 20분간 놓아두었으며, 이후 원심분리기(Hanil, Korea)를 사용하여 4,000 rpm, 4°C에서 20분간 원심분리하였다. 상등액은 0.45 µm 및 0.2 µm syringe filter(Advantec, Japan)를 순차적으로 이용하여 여과하였으며, 여과된 무세포 배양 상징액(cell-free supernatant, CFS)은 –20°C에서 보관하였다. 이 CFS는 병원성 미생물에 대한 생육 억제 활성 평가에 사용되었다. 또한, 동치미 유래 유산균의 CFS가 열 처리 후에도 항균 활성을 유지하는지를 확인하기 위해 100°C에서 5분간 열처리하였으며, 중성 조건에서의 항균 활성 변화를 확인하기 위하여 1N NaOH를 사용하여 pH를 7.0으로 조절한 뒤 항균 활성을 평가하였다.
2.5. 항균 활성 측정
동치미 유래 유산균의 배양 상징액이 병원성 세균의 생육을 저해하는지를 평가하기 위해 paper disc 확산법을 이용하였다. LB agar(0.7% agar) 20 mL를 제조하여 멸균한 후, 항온수조에서 50°C까지 냉각하였다. 이후 각 E. coli 균주 100 µL를 LB agar에 접종하고 고르게 혼합한 뒤, petri dish에 부어 고화시켰다. 배지가 완전히 굳은 후, 멸균된 paper disc를 배지 표면 위에 올리고, 각각 동치미 유래 유산균의 배양 상징액, 100°C에서 5분간 열처리한 상징액, 그리고 pH 7.0으로 조절한 상징액을 각 50 µL씩 disc에 흡수시켰다. 그 후 37°C 배양기에서 배양하여 병원성 세균의 생육 억제 여부를 관찰하였다. 한편, 항균 효과가 박테리오신 또는 유사 항균 펩타이드에 기인하는지를 확인하기 위하여, 배양 상징액을 Centriprep YM-5(Millipore, USA)를 이용해 분자량 5,000 Da 이상과 이하의 두 부분으로 분획하고, 각 분획물에 대한 항균 활성을 측정하였다.
2.6. SDS-PAGE와 Silver Staining
SDS-PAGE는 Laemmli [12]의 방법에 따라 12%의 gel로 수행하였다. 전기영동이 완료된 겔은 GE Healthcare(EU)에서 제공하는 silver staining kit를 사용하여 은염색을 실시하였다. 모든 과정은 제조사에서 제공하는 프로토콜에 따라 수행되었다. 겔 고정(fixation)은 100 mL의 ethanol과 25 mL의 glacial acetic acid를 혼합한 후 Milli-Q water로 총량을 250 mL로 맞춘 용액에 겔을 넣고, 30분간 천천히 진탕하여 수행하였다. Sensitizing은 ethanol 75 mL, 5% (w/v) sodium thiosulfate 10 mL, sodium acetate 17 g을 혼합한 뒤 Milli-Q water를 첨가하여 250 mL로 정량한 용액을 사용하여 15분간 2회 반복하여 진탕하였다. 감작이 완료된 겔은 Milli-Q water로 5분간 3회 세척하였다. Silver reaction은 2.5% (w/v) silver nitrate 용액 25 mL를 Milli-Q water로 250 mL로 희석한 후, 해당 용액의 절반을 겔에 첨가하여 20분간 천천히 진탕하였으며, 반응 도중 나머지 silver 용액을 추가로 부어 반응을 완료하였다. 밴드 현출(developing)은 sodium carbonate 6.25 g과 37% (w/v) formaldehyde 0.05 mL를 혼합하고 Milli-Q water로 250 mL까지 정량한 용액을 겔에 첨가하여, 밴드가 명확히 나타날 때까지 진탕하면서 진행하였다. 반응 종료(stopping)는 EDTA-Na₂·2H₂O 3.65 g을 Milli-Q water로 용해하여 250 mL로 맞춘 정지 용액을 사용하였으며, 이 용액에 겔을 넣고 10분간 진탕하여 반응을 중단하였다. 이후, 겔은 Milli-Q water로 5분씩 3회 반복하여 세정하였다.
3. Results and Discussion
3.1. 유산균의 분리 및 동정
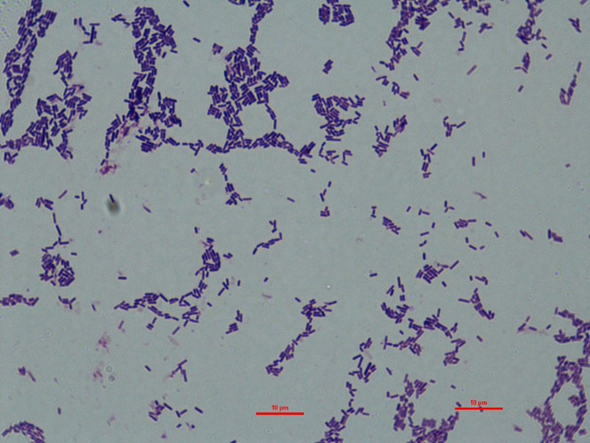
동치미로부터 유산균을 분리·동정하기 위해 sodium azide가 첨가된 MRS agar에 도말(streaking)한 후, 37°C 배양기에서 콜로니가 형성될 때까지 혐기성 조건에서 배양하였다. 배양이 완료된 후 선택된 콜로니에 대해 그람염색을 실시하여 세포 형태 및 그람 양성 여부를 확인하였다. 그 결과, 선택된 균주는 그람 양성의 간상형 세포로 확인되었다(Fig. 1). 또한, API 50 CHL 키트(bioMérieux, France)를 사용하여 당 이용성 분석을 수행한 결과, 해당 균주는 Lactobacillus plantarum과 99.9%의 상동성을 나타내었다(Table 1).
Table 1.
Carbohydrate utilization profile of Lactobacillus plantarum isolated from Dongchimi, as determined by the API 50 CHL kit
3.2. 동치미 유래 유산균의 항균 활성
동치미로부터 분리·동정된 L. plantarum의 항균 활성을 확인하기 위하여, 해당 균주를 MRS broth에 접종한 뒤 37°C 혐기 조건에서 24시간 배양하였다. 배양이 완료된 후, 배양 상징액은 0.2 µm syringe filter를 이용하여 여과하였으며, 이때 얻어진 상징액의 pH는 3.76이었다. 본 실험에서는 이 배양 상징액이 병원성 대장균 (E. coli)에 대해 항균 활성을 가지는지, 생성된 항균 물질이 열에 대해 안정한지, 그리고 중성 pH 조건에서도 활성이 유지되는지를 평가하였다. 관련 실험 결과는 Fig. 2에 제시하였다. 시험에 사용된 5종의 E. coli 균주—KCCM 11569(Fig. 2-A1), KCCM 11591(Fig. 2-B1), KCCM 11596(Fig. 2-C1), KCCM 11600(Fig. 2-D1), KCCM 11587(Fig. 2-E1)—에 대해 배양 상징액은 모두에서 뚜렷한 항균 환(clear zone)을 형성하며 강한 항균 활성을 나타내었다. 또한, 항균 물질의 열 안정성을 확인하기 위하여 배양 상징액을 100°C에서 5분간 열처리한 후 동일한 실험을 수행한 결과, 열처리 이전과 유사한 수준의 항균 환이 관찰되었으며(Fig. 2-A3~E3), 이는 해당 물질이 비교적 높은 열 안정성을 가지고 있음을 시사한다. 이러한 특성은 열처리 공정이 포함된 식품 가공 분야에서의 응용 가능성을 뒷받침한다. 한편, 배양 상징액의 pH를 1N NaOH를 사용하여 7.0으로 조절한 뒤 항균 활성을 측정한 결과, 모든 E. coli 균주(Fig. 2-A2~E2)에서 항균 환이 형성되지 않았다. 이는 L. plantarum이 생성한 항균 물질이 pH 변화, 특히 중성 또는 알칼리성 조건에서는 활성을 상실하며, 산성 환경에서만 안정적인 항균 활성을 유지함을 의미한다. 이러한 결과는 해당 물질이 pH 변화에 매우 민감하게 반응함을 보여주며, 그 특성을 보다 명확히 규명하기 위한 추가적인 연구가 필요함을 시사한다.

Fig. 2.
Antimicrobial activity of the cell-free supernatant produced by Lactobacillus plantarum isolated from Dongchimi against various Escherichia colistrains. A; E. coliKCCM 11569, B; E. coliKCCM 11591, C; E. coli KCCM 11596, D; E. coliKCCM 11600, E; E. coli KCCM 1187. 1; 50 µL untreated supernatant, 2; 50 µL supernatant adjusted to pH 7.0, 3; 50 µL heat-treated supernatant(100°C for 5 min).
3.3. 분자량별 상징액의 항균 활성 분석
동치미 유래 L. plantarum의 배양 상징액이 병원성 세균에 대해 나타내는 항균 활성이 박테리오신 또는 유사 항균 펩타이드에 기인하는지를 확인하기 위하여, Centriprep YM-5(Millipore, USA)를 이용하여 해당 상징액을 분자량 5,000 Da 이상 및 이하의 두 분획으로 나누었다. 이후 각 분획에 대해 Escherichia coli에 대한 항균 활성을 평가하였다. 실험 결과는 Fig. 3에 제시된 바와 같이, 두 분획 모두에서 병원성 균주의 생육을 억제하는 뚜렷한 항균 효과가 관찰되었다. 이는 L. plantarum이 생성한 항균 물질이 다양한 분자량 범위에 존재하며, 박테리오신 혹은 유사 항균 펩타이드의 복합적인 작용 가능성을 시사한다.

3.4. 전기영동을 통한 항균활성 분자량 확인
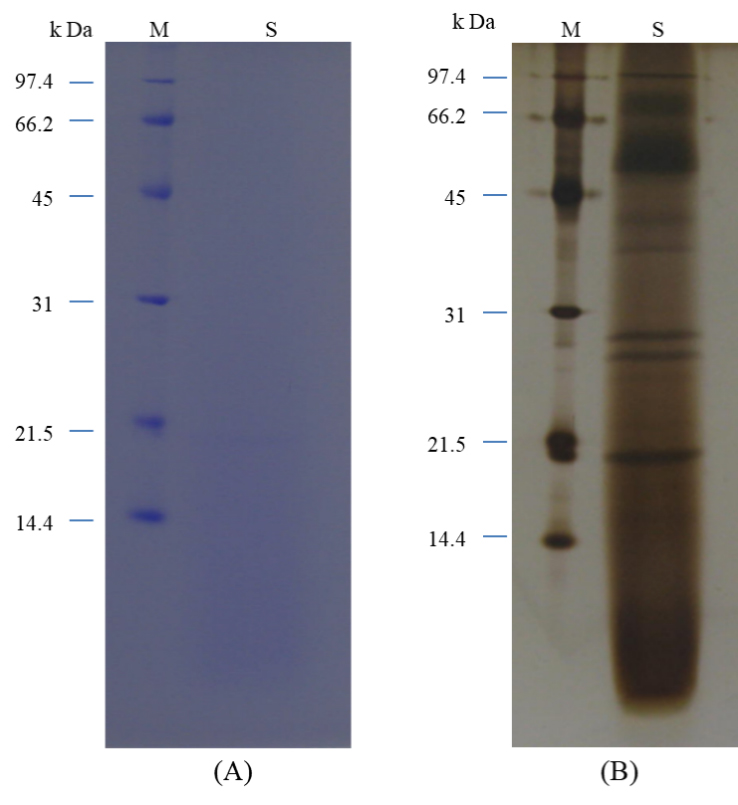
동치미 유래 L. plantarum의 배양 상징액에 포함된 항균 관련 물질의 분자량을 확인하기 위해 SDS-PAGE 분석을 수행하였다. Coomassie Brilliant Blue(CBB) 염색을 통한 밴드 관찰 결과(Fig. 4A), 전체 단백질 농도가 낮아 명확한 밴드의 시각적 확인이 어려웠다. 이에 민감도를 높이기 위해 silver staining을 적용한 결과(Fig. 4B), 여러 개의 단백질 밴드가 비교적 선명하게 관찰되었다. 그러나 관찰된 모든 밴드가 항균 활성을 가지는 것은 아니므로, 항균 활성을 갖는 특정 밴드를 확인하기 위한 후속 실험을 실시하였다. SDS-PAGE 후, 겔을 멸균 증류수로 세척하여 단백질 잔류물을 제거한 뒤, 병원성 세균이 포함된 agar 배지 위에 겔을 직접 올려 항균 활성을 평가하였다. 하지만 이 방법을 통해 항균 활성을 나타내는 특정 밴드를 명확히 식별할 수는 없었다(data not shown). 이러한 결과는 박테리오신 등 항균 물질의 정확한 위치 및 분자량 확인을 위해서는 L. plantarum의 배양 조건, 배양 시간, 상징액의 농축 과정 등을 고려한 추가적인 정제 및 확인 실험이 필요함을 시사한다.
유산균은 다양한 생리활성 물질을 생성하며, 이들 중 일부는 병원성 미생물의 생육을 억제하는 단백질성 항균 펩타이드인 박테리오신(bacteriocin)을 포함한다. 특히, 전통 발효식품으로부터 분리된 Lactobacillus 및 Pediococcus 종은 biogenic amine의 생성을 억제하는 활성을 보인 바 있으며[13], Lactococcus lactis subsp. lactis VR84 및 EG49는 nisin Z 및 lacticin 481을 생산하여 히스타민 생성균의 성장을 효과적으로 억제할 수 있음이 보고되었다[14]. 또한, Shin et al. [15]은 전통 발효식품으로부터 분리된 Lactobacillus salivarius LCH1227 균주가 Listeria monocytogenes에 대해 강한 항균 활성을 가지며, 이 균주가 생성한 항균 물질은 약 5 kDa의 분자량을 가진 단백질성 물질로 밝혀진 바 있다. 박테리오신은 일반적으로 단백질 또는 펩타이드 구조를 가지며, 인체 내에서는 단백질 분해효소에 의해 쉽게 분해되므로 잔류성이 없고 독성 또한 나타나지 않아 안전한 식품 보존제로서 주목받고 있다[16]. Lactobacillus 속 유산균이 생산하는 박테리오신의 대부분은 Class II형에 속하며, 저분자이며 열 안정성이 높아 식품 산업에서의 활용 가능성이 크다. 예를 들어, L. acidophilus, L. gasseri, L. johnsonii 등에서 박테리오신 생성이 보고되었으며, 이 중 대표적인 예인 Lactacin F는 후속 분류 연구를 통해 L. johnsonii에서 유래한 것으로 재정립되었다[17, 18, 19, 20]. 유산균은 인간뿐 아니라 가축의 장 건강 증진에도 긍정적인 영향을 미치는 것으로 알려져 있으며, 유기산, 과산화수소, 디아세틸, 박테리오신 등 다양한 항균 물질을 생성한다. 이 중 박테리오신은 미생물 기원 단백질성 항균 화합물로, 식품 보존의 천연 대체물질로서 주목받고 있다[21]. 유산균이 생산하는 박테리오신은 열에 강하고 산성 조건에서 안정하기 때문에, 가열 공정을 포함하는 식품 제조 환경에서도 적용이 가능하다. 대표적으로 Lactococcus lactis가 생성하는 nisin은 현재 50개국 이상에서 식품 첨가물로 사용이 승인되어 있으며, 이는 유산균 유래 항균 물질의 산업적 적용 가능성을 강하게 시사한다. 한편, 항생제 내성 병원균의 확산은 식품 안전과 공중보건에 심각한 위협이 되고 있으며, 이에 따라 소비자들은 저온살균 또는 무첨가 등 자연친화적 식품에 대한 선호를 보이고 있다. 그러나 저온처리만으로는 병원성 미생물의 완전한 제어가 어려우므로, 유산균 유래 항균 물질을 식품 보존제로 활용하는 것은 식중독 예방 및 식품 안전성 향상에 효과적인 전략이 될 수 있다. 또한, 유산균이나 그 유래 박테리오신을 가축 사료에 첨가할 경우, 장내 미생물 균형 개선과 질병 예방 효과를 기대할 수 있으며, 이는 유산균이 식품 산업뿐 아니라 축산 분야에서도 스타터 컬처 및 사일리지 품질 향상을 위한 유용 미생물제로 활용될 수 있음을 의미한다.
본 연구 결과, 동치미로부터 분리된 L. plantarum이 생성한 배양 상징액이 병원성 E. coli에 대해 우수한 항균 활성을 나타냄을 확인하였다. 그러나 본 연구에서는 해당 항균 활성이 박테리오신에 의한 것인지에 대한 직접적인 증거를 확보하지는 못하였으며, 항균 성분의 동정 및 구조적 특성에 대한 후속 연구가 필요하다. 이를 통해 유산균 유래 항균 물질의 분자적 메커니즘과 식품 보존제로서의 활용 가능성을 보다 명확히 규명할 수 있을 것이다.
4. Conclusions
유산균은 박테리오신을 포함한 다양한 항균 물질을 생성하며, 이는 병원성 미생물 억제 및 식품 보존에 효과적으로 활용될 수 있다. 특히 전통 발효식품에서 분리된 Lactobacillus 속 유산균은 항균 활성이 우수하며, 본 연구에서 동치미 유래 L. plantarum이 생성한 배양 상징액은 E. coli에 대해 강한 항균 활성을 나타냈다. 이 항균 물질은 열 안정성을 가지나, 중성 pH 조건에서는 활성을 상실하였다. 또한 분자량 분획 결과, 5 kDa 이상 및 이하 모두에서 항균 활성이 확인되어 복합적인 항균 인자의 존재 가능성을 시사하였다. 그러나 본 연구에서는 박테리오신 존재에 대한 직접적인 증거는 확보하지 못하였으며, 향후 항균 성분의 분리, 동정 및 구조 분석을 통해 그 작용 기전을 규명할 필요가 있다.
