

1. Introduction
2. Materials and Methods
2.1. Isolation and identification of lactic acid bacteria
2.2. Preparation of cell-free supernatant (CFS)
2.3. Bacterial strains
2.4. Hemolytic activity
2.5. Antibacterial activity of CFS derived from the isolated LAB
Paper disc diffusion assay
96 well plate assay
2.6. Statistical analysis
3. Results and Discussion
3.1. Isolation of lactic acid bacteria from Cynara scolymus L.
3.2. Identification of lactic acid bacteria isolated from Cynara scolymus L.
3.3. Antibacterial activity of cell-free supernatant (CFS) in different concentrations by paper disc method
3.4. Antibacterial activity of cell-free supernatant (CFS) in different concentrations by 96-well plate method
3.5. Hemolytic Activity of Lactobacillus plantarum
4. Conclusion
1. Introduction
According to the World Health Organization, access to safe and nutritious food is essential for maintaining health, preventing disease, and supporting sustainable development worldwide [1,2]. In this context, increasing attention has been directed toward functional foods and their bioactive components, including probiotics, prebiotics, and postbiotics, because of their demonstrated ability to modulate the gut microbiota, enhance immune responses, and improve overall health outcomes [3]. Among these components, probiotic microorganisms have become particularly important in the development of functional foods due to their capacity to confer health benefits when administered in adequate amounts [4]. The exploration of novel probiotic microorganisms from natural environments remains a major research focus. Microorganisms isolated from plants, soil, fermented foods, and animal-derived products represent valuable biological resources for the discovery of strains possessing unique physiological and functional characteristics [5,6]. Such strains may exhibit desirable probiotic traits, including tolerance to gastrointestinal conditions, antimicrobial activity against pathogenic microorganisms, antioxidant capacity, and the production of beneficial metabolites such as bacteriocins and postbiotic compounds [3,5]. The identification and characterization of these microorganisms provide opportunities for developing innovative fermented foods, natural preservatives, and next-generation probiotic products.
Artichoke (Cynara scolymus L.), originally from the warm regions of North Africa and the Mediterranean, has been successfully domesticated by Mongolian researchers to adapt to the harsh continental climate [7,8]. Among plant-derived sources, artichoke (Cynara scolymus L.) has emerged as a promising substrate for the isolation of beneficial microorganisms because it contains abundant bioactive compounds, including inulin, phenolic acids, flavonoids, and cynarin [9,10]. Inulin, a well-established prebiotic carbohydrate, selectively stimulates the growth of beneficial gut microorganisms, particularly lactic acid bacteria, while phenolic compounds contribute antioxidant and antimicrobial activities that may influence microbial ecology and functionality [10]. These characteristics suggest that artichoke may serve not only as a source of prebiotic compounds but also as a natural reservoir of probiotic microorganisms with potential applications in food biotechnology and human health. Although several studies have investigated the nutritional and prebiotic properties of artichoke, comparatively limited information is available regarding the isolation, physiological characterization, and functional evaluation of lactic acid bacteria naturally associated with this plant [9].
Therefore, the present study aimed to isolate and identify lactic acid bacteria from artichoke and to evaluate their probiotic and functional properties. Specifically, the study investigated whether artichoke-derived isolates possess characteristics desirable for probiotic applications, including tolerance to gastrointestinal stress conditions, antimicrobial activity against pathogenic bacteria, antioxidant potential, and other functional attributes relevant to food and health applications. We hypothesized that the prebiotic-rich environment of artichoke supports the growth of lactic acid bacteria exhibiting significant probiotic and bio functional properties. The findings of this study contribute to understanding the probiotic potential of artichoke-associated microorganisms and provide a scientific basis for their future utilization in functional food development, natural antimicrobial production, and fermentation technologies.
2. Materials and Methods
2.1. Isolation and identification of lactic acid bacteria
The fresh leaves of artichoke plants cultivated in a greenhouse in Ulaanbaatar were harvested in August 2024 and fermented under anaerobic conditions at 37°C for 4 weeks to enrich lactic acid bacteria (LAB). Following fermentation, the leaves were homogenized with sterile distilled water, and the resulting suspension was spread onto De Man Rogosa and Sharpe (MRS) agar (Difco, USA) supplemented with 0.01% sodium azide (Sigma-Aldrich, USA). The plates were incubated under both aerobic and anaerobic conditions at 37°C until visible colonies developed. Colonies were selected based on morphological characteristics, such as shape and size, and subjected to Gram staining and microscopic examination for preliminary identification. Biochemical characterization was performed using the API 50 CHL system (BioMérieux, Marcy-l'Étoile, France). Briefly, 100 µL of CHL medium inoculated with a single colony was dispensed into each capsule of the API strip and incubated at 37°C for 48 h. Carbohydrate fermentation profiles were recorded based on color changes and interpreted using API Web software (https://apiweb.biomerieux.com/) to determine strain identity. For molecular identification, 16S rRNA gene sequencing was outsourced to a commercial service provider (Solgent, Korea). The obtained sequence was compared to reference sequences in the NCBI database using BLAST analysis.
2.2. Preparation of cell-free supernatant (CFS)
The isolated LAB was sub-cultured in MRS broth and incubated aerobically at 37°C for 24 h. Following incubation, the culture was centrifuged at 4,000 rpm for 20 min at 4°C using a refrigerated centrifuge (Mega-17R, Hanil Science Industrial, Korea) to remove bacterial cells. The supernatant was collected and passed through a sterile 0.22 µm syringe filter (Advantec, Tokyo, Japan) to obtain a sterile CFS. The filtered CFS was aliquoted and stored at –20°C until further use in antibacterial and cellular bioactivity assays.
2.3. Bacterial strains
Four pathogenic strains (Salmonella 12021, Staphylococcus epidermidis, Enterococcus casseliflavus, Enterococcus faecalis) were obtained from the Korea Center for Microbial Protection and all strains were cultured in Luria-Bertani (LB) broth (Difco, USA). All strains were incubated at 37°C for 18 hours in an incubator to ensure active growth for antibacterial activity assays.
2.4. Hemolytic activity
The LAB isolate was pre-cultured in MRS broth at 37°C for 24 h and streaked onto blood agar plates containing 5% defibrinated sheep blood (Kisan Biotech Co., Seoul, Korea). The plates were incubated aerobically at 37°C for 48 h. Hemolytic activity was determined by examining zones of hemolysis around the colonies.
2.5. Antibacterial activity of CFS derived from the isolated LAB
Paper disc diffusion assay
The antibacterial activity of cell-free supernatant (CFS) obtained from isolated Lactobacillus plantarum against four pathogenic bacteria was evaluated using the paper disc diffusion method. LB (Luria–Bertani) agar (20 mL per plate) was sterilized at 121°C for 20 minutes and cooled to approximately 50°C. Each pathogenic strain (Salmonella 12021, Staphylococcus epidermidis, Enterococcus casseliflavus, and Enterococcus faecalis) was inoculated into the cooled LB agar at a concentration of 1% (v/v), then poured into sterile Petri dishes. After solidification, sterile paper discs (Advantec, Japan) were placed on the surface of each agar plate. The CFS obtained from L. plantarum was tested under three conditions:
(a) untreated CFS, (b) pH-adjusted pH 7.0 CFS, and (c) heat-treated CFS (100°C for 5 minutes). Each treatment (50 µL/disc) was absorbed onto the paper discs, and the plates were incubated at 37°C for 3 hours. Antibacterial activity was determined by measuring the diameter of the inhibition zones (mm) around each disc.
96 well plate assay
The antibacterial activity of CFS obtained from isolated L. plantarum against four pathogenic bacteria at each concentration was measured using the 96-well plate method. The prepared supernatant was added at concentrations of 20%, 10%, 5%, and 2.5% (v/v). After pre-culturing, 1% of each pathogenic bacterium was inoculated. The growth of pathogenic bacteria was cultured in an incubator at 37°C, and the optical density was measured at 655 nm wavelength using a microplate reader (Bio-Rad) at 3 h intervals.
2.6. Statistical analysis
All experimental data were analyzed using Microsoft Excel (Office 365, Microsoft Corp, WA, USA). Results were expressed as mean ± standard deviation (SD) based on three independent replicates.
3. Results and Discussion
3.1. Isolation of lactic acid bacteria from Cynara scolymus L.
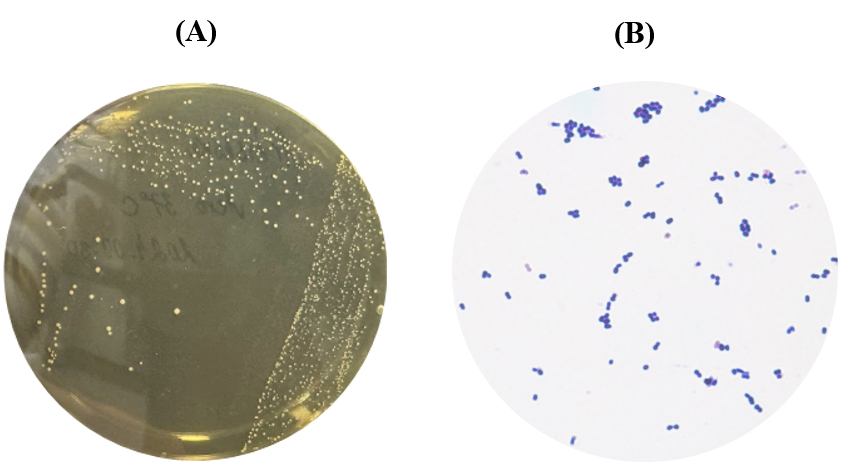
Fermented Cynara scolymus L. samples were homogenized with sterile distilled water and processed using a stomacher to obtain a uniform suspension. Appropriate dilutions were spread onto de Man, Rogosa and Sharpe (MRS) agar plates and incubated under both aerobic and anaerobic conditions at 37°C for 48 h [11]. Colonies exhibiting morphological characteristics typical of lactic acid bacteria (LAB) were selected and purified through repeated streaking. Preliminary identification was performed based on colony morphology, Gram staining, and microscopic observation. The selected isolates appeared as Gram-positive, non-spore-forming rod-shaped bacteria, which are characteristic features of the genus Lactobacillus[12]. Microscopic examination confirmed uniform rod morphology without evidence of contamination by cocci or Gram-negative bacteria (Fig. 1-A, B). The isolation of LAB from fermented artichoke demonstrates that C. scolymus provides a favorable ecological niche for the development of beneficial microorganisms during spontaneous fermentation. Previous studies have reported that vegetables and plant-derived fermented products harbor diverse LAB populations capable of producing bioactive metabolites with probiotic and antimicrobial functions [13,14]. Therefore, fermented artichoke may represent a valuable source of functionally important LAB strains for food and health-related applications.
3.2. Identification of lactic acid bacteria isolated from Cynara scolymus L.
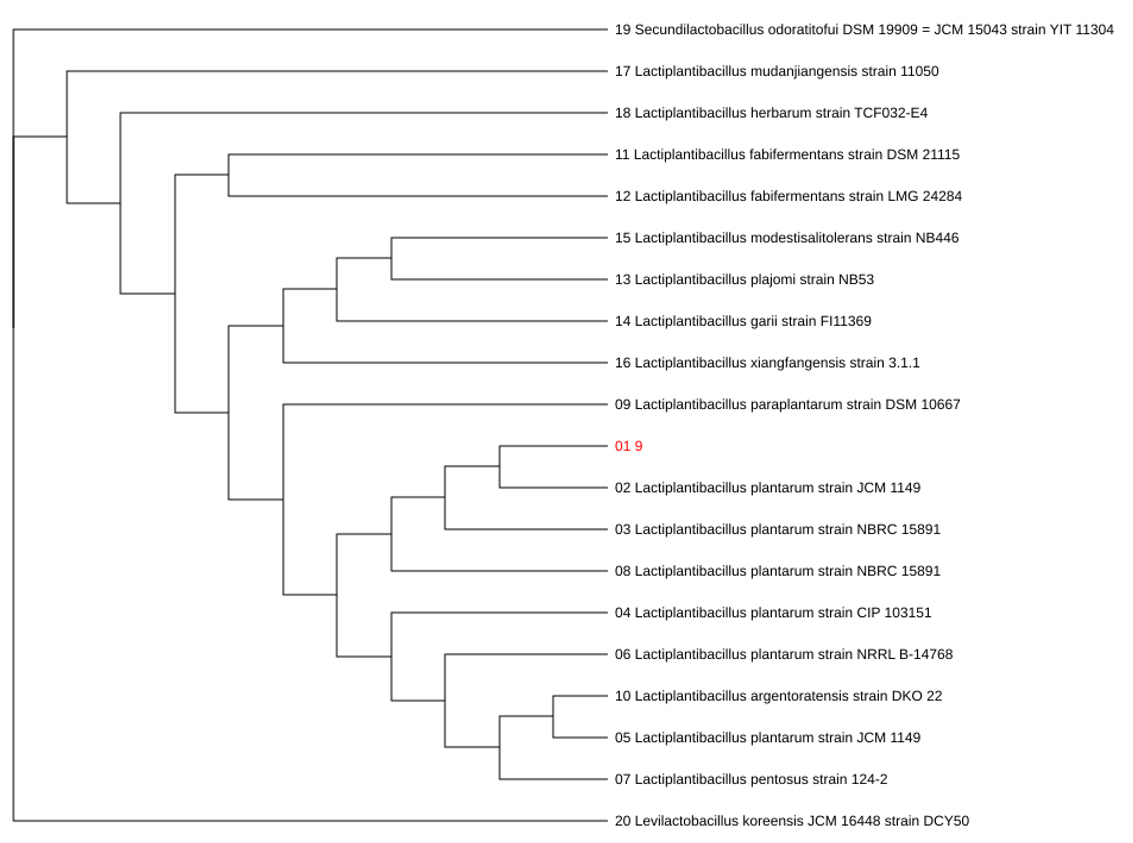
The selected isolate was initially characterized using the API 50 CHL carbohydrate fermentation system and subsequently confirmed through 16S rRNA gene sequencing. The carbohydrate utilization profile obtained from the API 50 CHL test is presented in Table 1, whereas the phylogenetic analysis based on 16S rRNA sequences is shown in (Fig. 2). The isolate fermented 28 of the 50 carbohydrates tested, including monosaccharides (L-arabinose, ribose, galactose, glucose, fructose, and mannose), disaccharides (maltose, lactose, sucrose, trehalose, and melibiose), sugar alcohols (mannitol and sorbitol), and several glycosides such as salicin, esculin, arbutin, amygdalin, and gentiobiose. In contrast, negative reactions were observed for glycerol, erythritol, D-arabinose, xylose isomers, dulcitol, inositol, xylitol, starch, glycogen, inulin, and ketogluconates. The fermentation profile corresponded closely to that reported for Lactobacillus plantarum, yielding a 99.9% identification probability. This result was further confirmed by 16S rRNA sequencing, which demonstrated 99.9% sequence similarity to Lactobacillus plantarum. The concordance between phenotypic and molecular analyses provides strong evidence for accurate taxonomic identification. The broad carbohydrate utilization spectrum observed in the present study reflects the remarkable metabolic versatility of L. plantarum. This species possesses an extensive repertoire of carbohydrate transport and catabolic genes, enabling adaptation to diverse ecological environments and fermentation substrates [15]. The ability to metabolize a wide range of plant-derived sugars is particularly advantageous in vegetable fermentation systems and contributes to its widespread application in food biotechnology and probiotic development [14,16]. Furthermore, the inability to ferment complex polysaccharides such as starch and inulin is consistent with previously reported metabolic characteristics of L. plantarum strains and can be used as an additional phenotypic marker for species differentiation [17].
Table 1.
Carbon utilization of the isolated strain Lactobacillus plantarium MULS using API 50CHL kit
3.3. Antibacterial activity of cell-free supernatant (CFS) in different concentrations by paper disc method
The antibacterial activity of the cell-free supernatant (CFS) produced by Lactobacillus plantarum isolated from Cynara scolymus was evaluated against four pathogenic bacteria using the agar disc diffusion method (Figs. 3, 4, 5, 6). Three treatments were examined, including untreated CFS (a), pH-adjusted CFS (pH 7.0) (b), and heat-treated CFS (100°C for 5 min) (c). Clear inhibition zones were observed around discs containing untreated CFS against Salmonella enteritidis, Staphylococcus epidermidis, Enterococcus faecalis, and Enterococcus casseliflavus, indicating that the isolate produced extracellular antimicrobial substances capable of suppressing both Gram-negative and Gram-positive bacteria. The inhibitory activity was markedly reduced or absent after neutralization to pH 7.0, suggesting that organic acids were major contributors to the observed antimicrobial effect. Heat treatment at 100°C for 5 min did not completely eliminate antibacterial activity in most cases, indicating that at least part of the antimicrobial activity was associated with heat-stable compounds. Such compounds may include organic acids and certain heat-resistant bacteriocin-like inhibitory substances that have previously been reported in L. plantarum strains [18,19]. Among the tested pathogens, Salmonella enteritidis and Staphylococcus epidermidis appeared to exhibit greater susceptibility to the untreated CFS, as evidenced by relatively larger inhibition zones, whereas Enterococcus casseliflavus showed comparatively weaker inhibition. Similar strain-dependent susceptibility patterns have been reported for LAB-derived antimicrobial metabolites [20]. These findings suggest that the antimicrobial activity of L. plantarum isolated from artichoke is primarily associated with acid production, although additional heat-stable antimicrobial metabolites may also contribute to pathogen inhibition. The ability to inhibit both Gram-positive and Gram-negative bacteria highlights the potential application of this strain as a natural bio-preservative and functional starter culture in food systems. The use of two different culture media served distinct experimental purposes. MRS broth was employed for cultivation of Lactobacillus plantarum and production of cell-free supernatant because it is specifically formulated to support the growth and metabolic activity of lactic acid bacteria. In contrast, Plate Count Agar was used as the assay medium for pathogenic bacteria because it provides uniform growth conditions and allows accurate visualization and measurement of inhibition zones. The separation of production and assay media minimizes potential interference from nutrients present in MRS medium and ensures a more reliable evaluation of antimicrobial activity.




3.4. Antibacterial activity of cell-free supernatant (CFS) in different concentrations by 96-well plate method
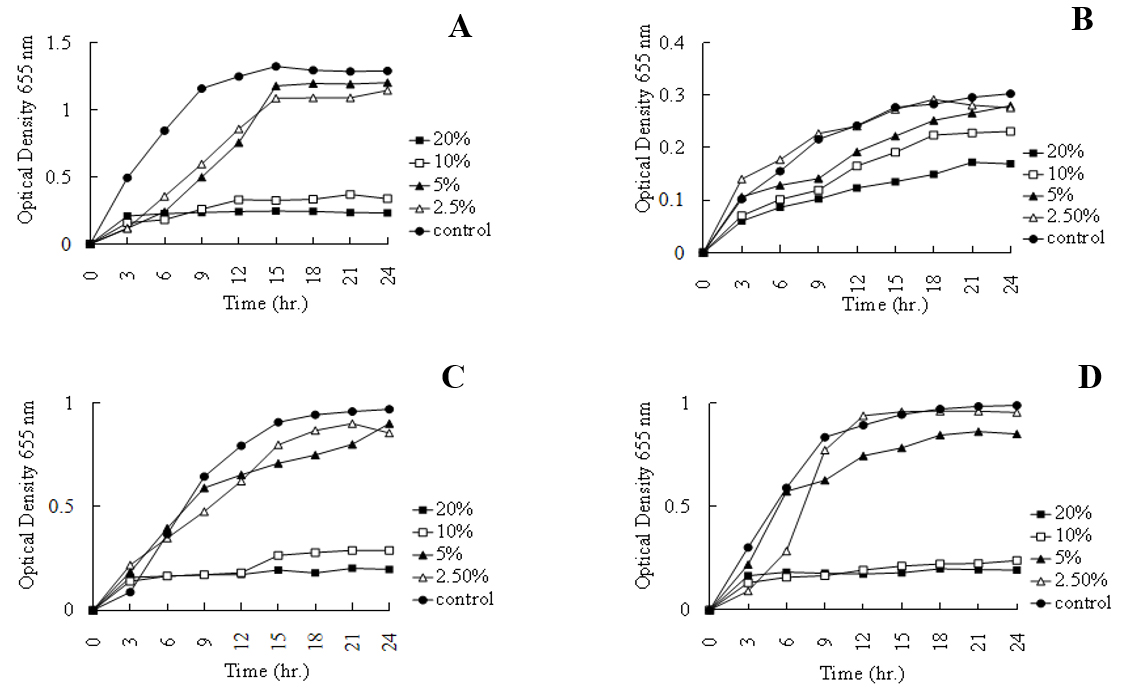
The antibacterial activity of L. plantarum CFS was further quantified using a 96-well microplate assay by monitoring bacterial growth through optical density measurements (OD600). The (Fig. 7) results demonstrated a clear concentration-dependent reduction in bacterial growth. Significant inhibition was observed at 20% and 10% CFS concentrations against Salmonella enteritidis, Staphylococcus epidermidis, and Enterococcus faecalis compared with untreated controls. In contrast, Enterococcus casseliflavus exhibited relatively greater resistance, maintaining measurable growth across all tested concentrations. The persistence of inhibitory activity even at 10% CFS, suggests the presence of highly active antimicrobial metabolites. Previous studies have shown that L. plantarum produces lactic acid, acetic acid, hydrogen peroxide, phenolic derivatives, and bacteriocin-like inhibitory substances that collectively suppress pathogen growth through membrane disruption, intracellular acidification, and metabolic inhibition [21,22]. The reduced inhibitory effect observed at 2.5% concentration likely reflects dilution of active compounds below the threshold required for effective growth suppression. These findings are consistent with previous reports demonstrating concentration-dependent antibacterial activity of LAB-derived metabolites against foodborne pathogens [23]. Overall, the strong inhibitory activity observed against both Gram-positive and Gram-negative bacteria highlights the potential of L. plantarum isolated from C. scolymus as a promising candidate for probiotic formulations and natural food preservation systems.
3.5. Hemolytic Activity of Lactobacillus plantarum
The safety of the isolated strain was evaluated using blood agar supplemented with 5% defibrinated sheep blood. No clear or greenish zones surrounding the colonies were observed after incubation, indicating a γ-hemolytic phenotype (Fig. 8). The absence of hemolysis suggests that the strain lacks hemolysins and other cytolytic factors capable of damaging erythrocyte membranes. Hemolytic activity is considered one of the most important safety indicators for probiotic microorganisms, and strains exhibiting γ-hemolysis are generally regarded as safe for food and human applications [24,25]. In contrast, α-hemolytic strains produce partial erythrocyte degradation, whereas β-hemolytic strains cause complete lysis of red blood cells and are often associated with pathogenicity and virulence [26]. Therefore, the non-hemolytic nature of the present isolate supports its safety profile and suitability for probiotic and food-related applications. The findings are consistent with previous reports indicating that L. plantarum possesses a long history of safe use in fermented foods and is generally recognized as a safe microorganism for industrial and probiotic applications [27].

4. Conclusion
This study successfully isolated lactic acid bacteria from fermented artichoke (Cynara scolymus L.) and identified the selected isolate as Lactobacillus plantarum through API 50 CHL analysis and 16S rRNA gene sequencing. The isolate demonstrated a broad carbohydrate utilization profile, reflecting its metabolic versatility and potential suitability for food fermentation applications. The cell-free supernatant (CFS) of L. plantarum exhibited significant antibacterial activity against several foodborne and opportunistic pathogens. The strongest inhibitory effects were detected against Salmonella enteritidis, Staphylococcus epidermidis, and Enterococcus faecalis, whereas Enterococcus casseliflavus exhibited comparatively lower susceptibility. These findings indicate that the isolate produces bioactive metabolites capable of suppressing the growth of both Gram-positive and Gram-negative pathogenic bacteria. Furthermore, the isolate displayed a γ-hemolytic phenotype on sheep blood agar, indicating the absence of hemolytic activity. No erythrocyte lysis or discoloration was observed around bacterial colonies, suggesting that the strain does not produce hemolysins or other cytotoxic factors associated with pathogenicity. This safety characteristic, together with its antimicrobial properties, supports the potential application of L. plantarum as a probiotic culture and natural bio-preservative in food systems. Future studies should focus on determining the minimum inhibitory concentration (MIC) of the antimicrobial metabolites, characterizing the active compounds responsible for pathogen inhibition, and evaluating the strain’s probiotic properties under simulated gastrointestinal conditions. In addition, the application of the isolated L. plantarum strain in fermented dairy products should be investigated to assess its technological performance, functional properties, and potential contribution to the development of locally produced probiotic and postbiotic foods that could reduce reliance on imported probiotic products.
