

1. Introduction
2. Materials and Methods
2.1. 공시동물 및 실험설계
2.2. 실험사료 및 사양관리
2.3. 조사항목 및 방법
2.4. 통계처리
3. Results and Discussion
3.1. 사양성적
3.2. 혈액 성상
3.3. 항산화 효과
4. Conclusions
1. Introduction
항영양인자는 사료 원료 내 존재하는 물질로 영양소 소화, 흡수 및 이용성을 저해하여 동물의 생산성 및 사료 가치를 저해하는 물질이며 이 중 oxalate는 항영양인자로 미네랄과 결합하여 대사를 방해하고 생산성을 저해한다고 알려져 있다[1, 2]. 호주, 뉴질랜드, 영국 등 양계 선진국에서는 동물복지 개선과 소비자의 요구 충족을 위해 약 40%의 농가가 방사 사육을 실시하고 있다[3]. 방사 사육 시 닭은 초지에서 자생하는 목초류를 자유롭게 섭취하는데, 일부 목초류에는 oxalate가 존재하며 특히 발육 초기에 가장 많은 oxalate를 축적한다[2, 4]. 체내에서 과도하게 축적된 oxalate는 칼슘(Ca)과 결합하여 신장결석을 유발할 수 있으며 이는 결과적으로 닭의 생산성을 저하한다고 알려져 있다[5].
Oxalate는 해당과정 중 pyruvate kinase의 작용을 억제하여 ATP 생성을 저해한다[6, 7]. 이로 인한 ATP 부족에 따른 에너지 대사 장애는 간 손상과 활성산소(ROS)의 생성, 세포 내 칼슘의 증가를 일으키며, 특히 세포 내 칼슘 증가는 미토콘드리아의 비선택적 물질 투과 채널 개방을 유도하여 이온과 작은 분자들의 자유로운 이동을 가능하게 해 막 전위를 손상시킴으로써 미토콘드리아의 팽창과 외막 파열을 유발한다[8]. 또한 체내 칼슘 증가와 같은 칼슘 불균형은 산화스트레스와 신장결석을 초래하며[9], 이러한 결과는 육계의 사양성적을 유의적으로 감소시키고 쇠약, 근육 위축, 신장 손상 및 폐사율 증가 등의 증상을 나타낸다[10].
Pyruvate는 에너지의 추가 공급 및 항산화제 역할을 수행한다고 알려져 있다[11]. 앞선 연구에 따르면 creatine pyruvate를 12 mg 투여 시 부화 후 육계 근육 내 ATP 농도가 0.31 µmol/g에서 0.40 µmol/g으로 유의적으로 증가하는 결과를 보였다[12]. 이는 pyruvate 보충이 ATP 추가 공급을 통해서 미토콘드리아 균형 유지와 에너지 공급을 지원할 수 있다는 것을 시사한다. Ramos-Ibeas et al. [13]은 인간 섬유아세포 및 배아줄기세포 배양 배지에 pyruvate를 2.5 mM 보충했을 때 H2O2를 효과적으로 제거함으로써 산화 스트레스를 억제했다고 보고하였으며, Long and Halliwell[14]은 pyruvate가 함유된 배지에서 H2O2가 효과적으로 제거되어 난소 세포 내 H2O2의 농도가 감소했다고 보고하였다.
현재 육계 사료 내 oxalate로 인한 항영양효과에 대한 pyruvate 첨가에 대한 연구는 제한적이다. 따라서, 본 연구는 oxalate의 항영양 효과를 완화하기 위한 pyruvate의 보충이 육계의 사양성적, 혈액성상, 항산화 활성에 미치는 영향을 평가하기 위해 수행하였다.
2. Materials and Methods
2.1. 공시동물 및 실험설계
본 연구는 University of Queensland의 동물실험윤리위원회의 승인을 받아(QAAFI/343/20), 본교의 가금류 연구 시설에서 수행하였다. 높은 수준의 oxalate 사료를 섭취한 육계에서 pyruvate의 보충이 사양성적, 혈액성상, 항산화 활성에 미치는 영향을 평가하기 위해 총 216마리의 1일령 육계(Ross 308, 42.13 ± 1.17)를 초기 체중을 기준으로 무작위로 대조구(CON), 1% oxalate 보충(Ox), 1% oxalate 및 1% pyruvate 보충(OxPyr)로 설계한 3가지 식이처리에 배치하였다. 각 처리군은 12반복, 반복당 6마리로 구성하였으며, 시험 기간은 Phase 1 (1‒14d), Phase 2 (15‒28d), Phase 3 (29‒42d)의 3단계로 나누어 총 42일간 실시하였다.
2.2. 실험사료 및 사양관리
실험사료의 영양소 함량은 Aviagen [15]에 제시된 영양소 요구량을 충족하거나 초과하도록 조성하였다(Table 1). 닭은 종이 깔짚이 깔린 평사 펜에서 사육하였으며, 사료와 물은 자유채식하도록 하였다. 계사 내 실내온도는 실험 시작 시 34°C로 설정하였으며, 8일차 이후 매주 3°C씩 점진적으로 낮추어 최종적으로 24°C에 도달하게 하였다. 모든 닭은 6일차까지 24시간동안 점등하였으며, 7일차부터 42일차까지는 16시간 점등, 8시간 소등으로 조정하였다.
Table 1.
Ingredients and nutrient composition of experimental diets, as-fed basis (Phase 1-3)
| Item | Day 1 to 14 | Day 15 to 28 | Day 29 to 42 | ||||||||
| CON | Ox | OxPyr | CON | Ox | OxPyr | CON | Ox | OxPyr | |||
| Ingredients, % | |||||||||||
| Maize-rolled | 62.27 | 58.31 | 58.17 | 65.69 | 60.94 | 60.43 | 66.93 | 62.29 | 61.91 | ||
| Soybean meal | 26.07 | 24.41 | 24.35 | 23.26 | 21.57 | 21.39 | 23.66 | 22.02 | 21.88 | ||
| MBM | 4.9 | 4.59 | 4.58 | 4.95 | 4.59 | 4.55 | 1.96 | 1.82 | 1.81 | ||
| Fish meal | 2 | 4.23 | 4.3 | 1 | 3.41 | 3.65 | 2 | 4.15 | 3.95 | ||
| Con-starch | - | 3 | 2.15 | - | 3.75 | 3.27 | - | 3.65 | 2.9 | ||
| Vegetable oil | 1.96 | 1.84 | 1.83 | 2.72 | 2.53 | 2.5 | 2.65 | 2.46 | 2.45 | ||
| Limestone | 1.08 | 1.01 | 1.01 | 0.99 | 0.92 | 0.91 | 1.37 | 1.28 | 1.27 | ||
| MDCP | 0.2 | 0.18 | 0.18 | - | - | - | 0.2 | 0.18 | 0.18 | ||
| Lysine HCL | 0.46 | 0.43 | 0.43 | 0.41 | 0.38 | 0.37 | 0.33 | 0.31 | 0.31 | ||
| DL-Methionine | 0.43 | 0.4 | 0.4 | 0.39 | 0.36 | 0.36 | 0.34 | 0.32 | 0.32 | ||
| L-Threonine | 0.2 | 0.18 | 0.18 | 0.16 | 0.15 | 0.15 | 0.12 | 0.11 | 0.11 | ||
| Salt | 0.2 | 0.18 | 0.18 | 0.2 | 0.18 | 0.18 | 0.2 | 0.18 | 0.18 | ||
| Mineral premix1) | 0.1 | 0.09 | 0.09 | 0.1 | 0.09 | 0.09 | 0.1 | 0.09 | 0.09 | ||
| Vitamin premix2) | 0.1 | 0.09 | 0.09 | 0.1 | 0.09 | 0.09 | 0.1 | 0.09 | 0.09 | ||
| Choline Chloride | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | ||
| Oxalate | - | 1 | 1 | - | 1 | 1 | - | 1 | 1 | ||
| Pyruvate | - | - | 1 | - | - | 1 | - | - | 1 | ||
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | ||
| Nutrient composition | |||||||||||
| ME, MJ/kg | 12.63 | 12.61 | 12.62 | 12.95 | 12.93 | 12.95 | 12.92 | 12.9 | 12.9 | ||
| CP, % | 22.23 | 22.23 | 22.23 | 20.55 | 20.55 | 20.54 | 19.81 | 19.81 | 19.81 | ||
| Lysine, % | 1.49 | 1.49 | 1.49 | 1.33 | 1.34 | 1.34 | 1.26 | 1.27 | 1.27 | ||
| Met+Cys, % | 1.11 | 1.09 | 1.09 | 1.02 | 1.01 | 1 | 0.98 | 0.96 | 0.96 | ||
| Threonine, % | 1.01 | 1 | 1 | 0.91 | 0.91 | 0.9 | 0.86 | 0.85 | 0.85 | ||
| Tryptophane, % | 0.24 | 0.24 | 0.24 | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 | ||
| Arginine, % | 1.38 | 1.38 | 1.38 | 1.27 | 1.27 | 1.27 | 1.21 | 1.21 | 1.22 | ||
| Calcium, % | 1.17 | 1.19 | 1.19 | 1.05 | 1.08 | 1.08 | 0.92 | 0.96 | 0.95 | ||
| P, % | 0.7 | 0.72 | 0.72 | 0.62 | 0.65 | 0.64 | 0.53 | 0.56 | 0.55 | ||
MDCP, Monodicalcium phosphate; Met+Cys, Methionine and cysteine; P, Phosphorouse; MBM, Meat and bone meal.
2.3. 조사항목 및 방법
사양성적은 14일차, 28일차, 42일차에 개체별로 체중과 사료 섭취량을 측정해 증체량 및 사료 요구율을 계산하였다.
혈액성상 분석과 항산화 분석을 위해 29일령에 펜당 1마리를 무작위로 선발하여 날개 정맥에서 혈액을 채취하여 즉시 15 ml Ethylene Diamine Tetraacetic Acid (EDTA) 튜브에 담았다. 채취한 혈액은 원심분리기에서 4°C에서 10분간 2000 g로 원심분리해 혈청을 분리하였고, 분석 전까지 –80°C에서 보관하였다.
혈액성상 분석을 위한 혈청 내 Alanine aminotransferase (ALT), Aspartate aminotransferase transaminase (AST), Alkaline Phosphatase (ALP), Gamma Glutamyl Transpeptidase (GGT), T- Bilirubin (T-Bil)의 수준은 채취한 혈청 3 ml를 사용하여 혈액생화학분석기(7180, HITACHI)를 이용하여 분석하였다.
혈청 내 cortisol 수준은 ELISA kit (Enzo Life Science, New York, USA)를 사용해 제조사의 지침에 따라 분석하였다. 혈청 내 항산화 효소 분석을 위해GSTs (Glutathione S Transferases), GSH-Px (Glutathione Peroxidase), Superoxide Dismutase (SOD), Total antioxidant capacity (TAOC)의 분석을 위해 ELISA kit (AFG Scientific, Illinois, USA)를 사용해 제조사의 지침에 따라 분석하였다.
2.4. 통계처리
본 실험에서 측정된 데이터의 유의성은 SAS 버전 9.4를 사용하였으며, 처리구 간 평균을 비교하기 위해 t-test를 이용하였다. p-value가 0.05 미만일 때 통계적으로 유의적인 차이가 나타나는 것으로 판단하였으며, 0.05 이상 0.1 미만일 때 경향이 나타나는 것으로 판단하였다.
3. Results and Discussion
3.1. 사양성적
육계 사료 내 oxalate의 항영양 효과를 완화하기 위한 pyruvate 보충이 사양성적에 미치는 영향은 Fig. 1과 같다. 본 연구 결과 BWG는 Phase 1‒3에서 대조군에 비해 oxalate를 1% 보충하여 급여한 Ox 처리구와 oxalate 1% 및 pyruvate 1%를 동시 보충 급여한 OxPyr 처리구에서 유의적으로 낮은 결과를 나타냈으며(p < 0.05), 전체 실험 기간에서는 Ox 처리구가 OxPyr 처리구와 비교했을 때 유의적으로 낮은 결과를 나타냈다(p < 0.05). FI는 전체 실험 기간동안 Ox 처리구와 OxPyr 처리구가 대조군과 비교했을 때 유의적으로 낮은 결과를 나타냈으며(p < 0.05), FCR은 처리구간 유의적인 차이가 나타나지 않았다(p > 0.05). Oxalate는 사료 작물에 함유되어 있는 항영양인자로 동물이 섭취했을 때 미네랄과 킬레이트 결합을 형성함으로써 체내 미네랄 흡수 및 대사를 저해하고 생산성을 감소시키며, 이러한 영향은 단위동물이 반추동물보다 민감하게 나타난다고 알려져 있다[2]. Koni et al. [16]은 oxalate가 47.8 mg/kg 함유된 사료를 급여한 육계에서 oxalate가 포함되지 않은 사료를 급여한 대조군에 비해서 BWG가 26.4 g/d, FI가 3.3 g/d 유의적으로 감소시켰다고 보고하였다. 또한 Robertson et al.[17]은 사료 내 oxalate의 함량을 0, 0.25, 0.50, 1%로 첨가하였을 때 6주령 육계의 체중이 각각 495, 464, 460, 392 g으로 감소했다고 보고하였다. 본 연구에서 Ox 처리구의 사양 성적 저하는 oxalate가 Mg2+, Mn2+ 등 성장에 필수적인 미네랄과 불용성 복합체를 형성하여 미네랄의 생체 이용률을 저하시는 데 기인한 것으로 판단된다. 특히 Mn의 생체이용률 감소는 사양 성적 저하와 밀접한 관련이 있는데, de Carvalho et al.[18]은 육계에서 Mn 공급이 부족할 경우 ADG가 2.12 g/d 만큼 유의적으로 감소했으며, FCR이 증가했다고 보고하였다.
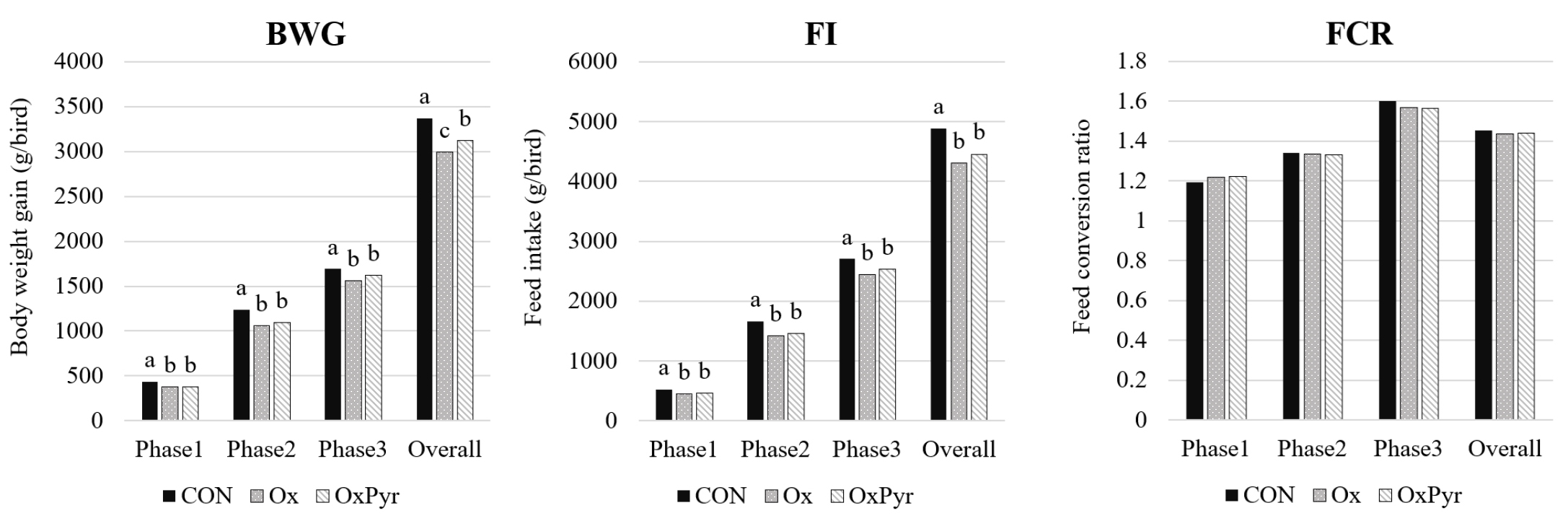
또한 oxalate는 해당과정의 pyruvate kinase 활성을 억제하여 ATP 생성 경로를 방해함으로써 에너지 대사를 저해할 수 있는 것으로 알려져 있다[7]. 외인성 pyruvate를 보충했을 때 pyruvate를 통한 추가적인 ATP 공급으로 oxalate에 의한 에너지 대사 저해 효과가 완화될 수 있다[12]. 본 연구결과에 있어 실험 전체 기간 동안 세 처리구간 BWG에서 유의적인 차이가 나타난 것은 oxalate에 의해 저해된 에너지 대사가 pyruvate 보충에 의해 부분적으로 개선되었기 때문으로 생각된다.
3.2. 혈액 성상
사료 내 oxalate와 pyruvate의 첨가에 따른 혈액 성상 결과는 Table 2와 같다. 본 연구 결과 AST 수치가 pyruvate 첨가에 따라 OxPyr에서 Ox에 비해서 유의적으로 감소하였고, cortisol은 oxalate 첨가에 따라 Ox에서 유의적으로 증가하였으며 OxPyr에서 대조군 수준으로 감소하였다(p < 0.05). ALT, ALP, GGT, T-Bil에서는 유의적인 차이가 발견되지 않았다(p > 0.05). AST는 미토콘드리아에 80%, 세포질에 20%가 분포하며 간 손상 지표로 주로 사용된다[19, 20]. Oxalate는 미토콘드리아 막 전위 균형의 불균형을 초래하여 미토콘트리아 팽창과 외막 파열을 유도함으로써 AST의 유리를 증가시켜 AST 농도를 증가시킨다[8]. Dong et al.[21]은 ethyl pyruvate가 돼지의 출혈성 쇼크로 인한 혈청 내 AST의 과잉 발현을 정상 수준으로 낮출 수 있다고 보고하였다. 또한 pyruvate는 미토콘드리아에 추가적인 ATP를 공급하여 미토콘드리아 기능의 안정화와 막 전위를 보호한다[22]. 이는 외인성 pyruvate의 추가적인 ATP 공급이 미토콘드리아 손상으로 인한 AST의 유리를 억제할 수 있음을 시사한다.
Table 2.
Effects of oxalate and pyruvate supplementation on serum biochemical indicators in broiler chickens (d 29)
| Group1) | CON | Ox | OxPyr | SEM2) | p-value |
| ALT (U/L) | 1.267 | 1.417 | 0.967 | 0.12 | 0.225 |
| AST (U/L) | 401ab | 456a | 296b | 27.52 | 0.032 |
| ALP (U/L) | 8329 | 9536 | 10783 | 848.51 | 0.336 |
| GGT (U/L) | 15.40 | 13.73 | 15.91 | 0.66 | 0.501 |
| T-Bil (mg/dL) | 0.927 | 0.912 | 0.957 | 0.03 | 0.794 |
| Cortisol (pg/ml) | 1895b | 2996a | 1626b | 210.30 | 0.004 |
Cortisol은 스트레스 환경에 노출되었을 때 증가하는 호르몬으로 장내 칼슘 흡수를 저하시키고, 지속적인 스트레스를 받은 동물에서 Ca-Oxalate (CaOx) 결석 형성을 촉진할 수 있다[23, 24, 25]. Oxalate가 미토콘드리아 에너지 대사 장애 및 ROS 생성을 유도하였고, cortisol은 이러한 미토콘드리아 산화스트레스의 직접적인 결과로 인해 증가된 것으로 사료된다[25]. 또한 Cao et al.[26]에 따르면 미토콘드리아의 oxalate 노출은 ROS의 축적을 증가시키고, 이로 인해 미토콘드리아의 기능적 장애를 일으킨다고 보고하였다. 본 연구에서 혈청 내 cortisol 수준이 감소한 것은 pyruvate가 항산화제로 작용해 oxalate로 인한 미토콘드리아의 산화 스트레스 완화에 기인한 것으로 생각된다[11, 27].
3.3. 항산화 효과
사료 내 oxalate와 pyruvate의 첨가에 따른 항산화 효과에 미치는 영향은 Table 3와 같다. 혈청 내 GSTs의 농도는 oxalate 첨가에 따라 유의적으로 감소하였고, SOD는 pyruvate 첨가에 따라 OxPyr에서 Ox에 비해 유의적으로 감소하였다(p < 0.05). GSH-Px와 T-AOC는 Ox 처리구에서 유의적으로 높게 나타났고 pyruvate 첨가에 따라 OxPyr 처리구에서 유의적으로 감소하였다(p < 0.05).
GSTs는 세포질 또는 미토콘드리아에 존재하며 세포 내 산화 환원 균형을 유지하는 데 기여하는 효소이다[28, 29, 30]. 그러나 GSTs는 ROS에 의해 불활성화 되며 GSTs가 가진 Glutathione 결합 부위가 산화에 의해 비가역적으로 손상될 수 있다[31]. Schroer et al.[32]은 마우스 모델에서 ROS로 인해 GST 종류 중 하나인 GSTPi의 활성이 감소했다고 보고하였다. 이는 oxalate로 유도된 ROS에 의해 GSTs 활성이 비가역적으로 저해되었음을 시사한다. 또한 Vanhaecke et al.[33]은 세포 배양액에 pyruvate를 첨가할 경우 GST 및 관련 단백질의 활성이 억제된다고 보고하였으며, 본 실험 결과도 이와 유사한 결과를 나타냈다.
Table 3.
Effects of oxalate and pyruvate supplementation on serum antioxidant concentrations and total antioxidant capacity in broiler chickens (d 29)
| Group1) | CON | Ox | OxPyr | SEM2) | p-value |
| GSTs (ng/ml) | 30.96a | 17.04b | 17.35b | 1.90 | 0.001 |
| GSH-Px (ng/ml) | 71.65b | 90.92a | 73.82b | 3.46 | 0.027 |
| SOD (U/L) | 499ab | 560a | 459b | 15.49 | 0.041 |
| T-AOC (U/ml) | 3.22b | 6.78a | 3.73b | 0.51 | 0.001 |
GSH-Px는 Glutathione을 사용해 H2O2 및 기타 과산화물을 환원시킴으로써 세포를 산화적 손상으로부터 보호하는 항산화 효소로, GSH-Px의 활성 증가는 산화 스트레스에 대한 반응성 증가를 나타내는 지표로 간주된다[34]. 이전 연구에서는 육계 사료에 3 mEq/kg의 지질 과산화물을 보충했을 때 GSH-Px 활성이 유의적으로 증가했다고 보고하였으며[35], H2O2로 유도된 세포사멸 연구에서는 pyruvate를 100 µM에서 4 mM까지 처리했을 때 세포 생존율이 용량 의존적으로 증가했다고 보고하였다[22]. 이는 oxalate로 유도된 ROS 생성이 산화 스트레스를 증가시켰고, 이를 완화하기 위해 항산화 효소 수준이 증가한 것으로 판단된다. OxPyr 처리구의 GSH-Px 수준이 유의적으로 감소한 결과는 pyruvate의 보충이 oxalate에 의해 생성된 H2O2을 제거함으로써 산화 스트레스 수준을 완화시켰기 때문으로 사료된다[36].
SOD는 슈퍼옥사이드 라디칼(O2⁻)을 H2O2로 전환시키는 항산화 효소로, ROS 제거의 초기 단계에서 핵심적인 역할을 수행하며[37], SOD의 작용 이후 항산화 효소들이 H2O2를 물과 산소로 분해하여 산화적 손상을 방지한다고 알려져 있다[38]. 본 연구 결과 Ox 처리구에서 SOD 수준이 유의적으로 높게 나타난 것은 oxalate에 의해 유도된 ROS를 제거하고, 특정 산화 조건에서 탈카르복실화 및 산화되는 과정에서 생성될 수 있는 O2⁻을 H2O2로 전환하기 위한 생리적 반응에 의해 효소 수준이 증가된 것으로 사료된다[39]. OxPyr 처리구의 SOD 활성은 유의적으로 낮게 나타났는데, 이는 pyruvate의 비효소 산화적 탈탄산 반응을 통해 H2O2를 직접 제거함으로써 산화스트레스를 완화시켜 SOD 활성이 상대적으로 감소한 결과로 판단된다[40].
T-AOC은 항산화 효소와 비효소계 항산화 물질을 모두 포함한 항산화 물질의 총량을 의미하며, 산화 스트레스 상태를 반영하는 지표로 사용된다[41]. 본 연구 결과 Ox에서 유의적으로 높게 나타난 것은 oxalate로 인한 산화 스트레스에 대응하기 위해 항산화 물질의 수준이 증가한 결과로 생각된다[42]. 반면 OxPyr 처리구에서는 T-AOC 수치가 유의적으로 낮게 나타났는데, 이는 oxalate에 의해 유도된 산화 스트레스가 pyruvate 처리에 의해 억제되어 방어 반응이 완화되었음을 나타내는 것으로 사료된다.
본 연구 결과 pyruvate의 첨가는 oxalate에 의한 육계의 사양성적, 혈액 성상 및 항산화 효과를 부분적으로 개선시킬 수 있음을 시사한다. 반면 육계 사료 내 oxalate의 항영양 효과를 완화하기 위한 pyruvate의 사용에 관한 연구는 부족한 실정으로, 이에 대한 추가적인 연구가 필요할 것으로 생각된다.
